Clasificación de los microorganismos según sus formas principales. Las bacterias, su diversidad. Estructura. Vitalidad. Términos básicos: bacterias
Nutrición de las bacterias.
Alimento.
Difusión pasiva
Difusión facilitada
transporte activo
En el primer caso, la molécula de nutriente forma un complejo con una proteína del espacio periplásmico que interactúa con una permeasa de membrana citoplasmática específica. Después de la penetración dependiente de la energía a través de la membrana citoplasmática, el complejo “permeasa de proteína periplasma-sustrato” se disocia con la liberación de la molécula de sustrato.
Durante el transporte activo con modificación química de la sustancia transportada, la cadena de eventos incluye: (1) fosforilación de la enzima de membrana 2 del citoplasma por fosfoenolpiruvato; (2) unión en la superficie de la membrana citoplásmica por moléculas de sustrato de enzima-2 fosforiladas; (3) transporte dependiente de energía de la molécula de sustrato al citoplasma; (4) transferencia de un grupo fosfato a una molécula de sustrato; (5) disociación del complejo "sustrato-enzima" en el citoplasma. Debido a la fosforilación, las moléculas de sustrato se acumulan en el citoplasma de las células y no pueden salir de ellas.
Clasificación de las bacterias por tipo de alimento.
A modo de ingesta de nutrientes. Las bacterias se clasifican en holófitos y Holozoico. bacterias holófitas (del griego. holos- completo y fiticos- relacionados con las plantas) no pueden liberar en el medio ambiente enzimas que descomponen los sustratos, por lo que consumen nutrientes exclusivamente en forma molecular disuelta. bacterias holozoicas (del griego. holos- completo y zoikos- relacionados con los animales), por el contrario, tienen un complejo de exoenzimas que proporcionan nutrición externa: la descomposición de sustratos en moléculas fuera de la célula bacteriana. Después de eso, las moléculas de nutrientes ingresan al interior de las bacterias holozoicas.
Por fuente de carbono aislado de bacterias autótrofos y heterótrofos. autótrofos (del griego. autos- mí mismo, trofeo- alimentos) el dióxido de carbono (CO 2) se utiliza como fuente de carbono, a partir del cual se sintetizan todas las sustancias que contienen carbono. Para heterótrofos (del griego geteros - otro y trofeo- alimentos) las fuentes de carbono son diversas sustancias orgánicas en forma molecular (carbohidratos, alcoholes polihídricos, aminoácidos, ácidos grasos). El mayor grado de heterotrofia es inherente a los procariotas, que solo pueden vivir dentro de otras células vivas (por ejemplo, rickettsia y clamidia).
Por fuente de nitrógeno los procariotas se dividen en 3 grupos: 1) bacterias fijadoras de nitrógeno (asimilan el nitrógeno molecular del aire atmosférico); 2) bacterias que consumen nitrógeno inorgánico de sales de amonio, nitritos o nitratos; 3) bacterias que asimilan el nitrógeno contenido en compuestos orgánicos (aminoácidos, purinas, pirimidinas, etc.).
Por fuente de energía Las bacterias se dividen en fotótrofos y quimiotrofos. bacterias fototróficas , como las plantas, son capaces de utilizar la energía solar. Los procariotas fototróficos no causan enfermedades en humanos. bacterias quimiotróficas reciben energía en reacciones redox.
Por la naturaleza de los donantes de electrones. litótrofos(del griego. litos- piedra) y organótrofos. A litótrofos (quimiolitotrofos ) las sustancias inorgánicas actúan como donantes de electrones (H 2, H 2 S, NH 3, azufre, CO, Fe 2+, etc.). Donadores de electrones organótrofos (quimioorganotrofos ) son compuestos orgánicos - carbohidratos, aminoácidos, etc.
La mayoría de las bacterias patógenas para los humanos tienen un tipo de nutrición quimioorganotrófica (quimioheterótrofa); el tipo quimiolitotrófico (quimioautotrófico) es menos común.
Por la capacidad de sintetizar compuestos orgánicos. Las bacterias quimiotróficas se clasifican en protótrofos, auxótrofos y hipotrofos. bacterias prototróficas sintetizar todas las sustancias orgánicas necesarias a partir de glucosa y sales de amonio. Las bacterias se llaman auxótrofos si no son capaces de sintetizar ninguna sustancia orgánica a partir de los compuestos indicados. El grado extremo de pérdida de la actividad metabólica se denomina desnutrición. bacterias hipotróficas proporcionan su actividad vital reorganizando las estructuras o metabolitos del huésped.
Además del carbono y el nitrógeno, el azufre, el fósforo y los iones metálicos son necesarios para la vida completa de las bacterias. Las fuentes de azufre son los aminoácidos (cisteína, metionina), vitaminas, cofactores (biotina, ácido lipoico, etc.), sulfatos. Las fuentes de fósforo son los ácidos nucleicos, los fosfolípidos, los fosfatos. En concentraciones suficientemente altas, las bacterias necesitan magnesio, potasio, calcio, hierro; en cantidades mucho más pequeñas: zinc, manganeso, sodio, molibdeno, cobre, níquel, cobalto.
factores de crecimiento- Estas son sustancias que las bacterias no pueden sintetizar por sí mismas, pero las necesitan con urgencia. Los aminoácidos, las bases nitrogenadas, las vitaminas, los ácidos grasos, las porfirinas de hierro y otros compuestos pueden actuar como factores de crecimiento. Para crear condiciones óptimas para la actividad vital de las bacterias, se deben agregar factores de crecimiento a los medios nutritivos.
Metabolismo, conversión de energía.
A) Metabolismo constructivo.
Una fase obligatoria de la nutrición bacteriana es la asimilación de nutrientes, es decir, su inclusión en forma alterada o modificada en reacciones sintéticas para la reproducción de componentes y estructuras celulares.
Metabolismo de proteínas en las bacterias, puede proceder en 3 fases: degradación primaria de proteínas, degradación secundaria de proteínas y síntesis de proteínas. La descomposición primaria de las moléculas de proteína en peptonas se lleva a cabo mediante exoenzimas, exoproteasas liberadas por bacterias en el medio ambiente. La descomposición secundaria ocurre bajo la acción de endoenzimas (endoproteasas), que tienen todas las bacterias. Este proceso tiene lugar dentro de la célula bacteriana y consiste en la descomposición de los péptidos en sus aminoácidos constituyentes. Estos últimos pueden utilizarse inalterados o ser sometidos a transformaciones químicas (desaminación, descarboxilación, etc.), que resultan en la aparición de amoníaco, indol, sulfuro de hidrógeno, cetoácidos, alcohol, dióxido de carbono, entre otros.La detección de estos compuestos es de importancia diagnóstica en bacteriología.
Junto con las reacciones de escisión de proteínas, ocurren reacciones de su síntesis. Algunas bacterias forman proteínas a partir de aminoácidos preparados obtenidos como resultado de la nutrición externa, mientras que otras bacterias sintetizan aminoácidos de forma independiente a partir de compuestos simples que contienen nitrógeno y carbono. La síntesis de aminoácidos puede llevarse a cabo en las reacciones de aminación, transaminación, amidación, carboxilación. La mayoría de los procariotas son capaces de sintetizar todos los aminoácidos que componen las proteínas celulares. Una característica de la biosíntesis de aminoácidos es el uso de rutas biosintéticas comunes: el ciclo del ácido tricarboxílico, la glucólisis, la ruta oxidativa de las pentosas fosfato, etc. El principal compuesto inicial para la síntesis de aminoácidos es el piruvato y el fumarato.
metabolismo de los carbohidratos difiere en autótrofos y heterótrofos (Esquema 1). Las bacterias autótrofas sintetizan todos los carbohidratos necesarios a partir del dióxido de carbono. Las materias primas para la formación de carbohidratos en bacterias heterótrofas pueden ser: (1) compuestos de uno, dos y tres carbonos; y (2) polisacáridos (almidón, glucógeno, celulosa). Para desdoblar estos últimos, muchas bacterias heterótrofas disponen de exoenzimas (amilasa, pectinasa, etc.), que hidrolizan polisacáridos para formar glucosa, maltosa, fructosa, etc.
En las bacterias autótrofas, en el ciclo de Calvin, se forma ribulosa fosfato-fósforo-ácido glicérico a partir del dióxido de carbono, que se incluye en las reacciones de glucólisis que van en sentido contrario. El producto final de la síntesis inversa es la glucosa.
Las bacterias heterótrofas forman glucosa a partir de compuestos de uno, dos y tres carbonos, incluyéndolos también en la reacción de glucólisis inversa. Debido al hecho de que algunas reacciones de glucólisis son irreversibles, los heterótrofos han formado reacciones enzimáticas especiales que les permiten eludir las reacciones irreversibles de la vía catabólica.
Cuando las bacterias heterótrofas escinden los polisacáridos, los disacáridos resultantes entran en las células y, bajo la influencia de la maltosa, la sacarosa y la lactosa, sufren hidrólisis y descomposición en monosacáridos, que luego se fermentan o se incluyen en las reacciones de interconversión de azúcar.
metabolismo de los lípidos. Tanto los lípidos exógenos como los anfibolitos del metabolismo intersticial pueden servir como materiales de partida para la formación de lípidos en bacterias. Los lípidos exógenos están expuestos a lipasas bacterianas y otras enzimas lipolíticas. Muchos tipos de bacterias absorben glicerol, que sirve como fuente de material plástico y energía. Las fuentes endógenas para la síntesis de lípidos pueden ser acetilcoenzima A, propionil-APB, malonil-APB (ACP - proteína transportadora de acetilo), fosfodioxiacetona, etc.
El sustrato inicial para la síntesis de ácidos grasos con número par de átomos de carbono es la acetilcoenzima A, para ácidos grasos con número impar de átomos de carbono, propionil-APB y malonil-APB. La formación de dobles enlaces en la molécula de ácido en procariotas aerobios ocurre con la participación del oxígeno molecular y la enzima desaturasa. En los procariotas anaerobios, los dobles enlaces se introducen temprano en la síntesis como resultado de una reacción de deshidratación. El sustrato inicial para la síntesis de fosfolípidos es la fosfodioxiacetona (un compuesto intermedio de la vía glucolítica), cuya reducción da lugar a la formación de 3-fosforoglicerol. Luego se le añaden 2 residuos de ácidos grasos en forma de complejo con APB. El producto de reacción es el ácido fosfatídico, cuya activación con CTP y posterior unión al grupo fosfato de serina, inositol, glicerol u otro compuesto conduce a la síntesis de los fosfolípidos correspondientes.
Los microorganismos que son auxotróficos e hipotróficos para los ácidos grasos (por ejemplo, los micoplasmas) los obtienen preparados a partir de células huésped o de un medio nutritivo.
Intercambio de mononucleótidos. Los mononucleótidos de purina y pirimidina son componentes esenciales del ADN y el ARN. Muchos procariotas son capaces tanto de utilizar bases de purina y pirimidina listas para usar, sus nucleósidos y nucleótidos contenidos en el medio nutritivo, como de sintetizarlos a partir de sustancias de bajo peso molecular. Las bacterias tienen enzimas que catalizan las siguientes etapas de interconversiones de derivados exógenos de purina y pirimidina: base nitrogenada - nucleósido - nucleótido (mono - di - - trifosfato).
Síntesis de mononucleótidos de purina y pirimidina. de novo llevado a cabo de manera independiente. Durante la síntesis de nucleótidos de purina, como resultado de sucesivas reacciones enzimáticas, se forma ácido inosínico, a partir del cual se sintetizan los ácidos adenilo (AMP) y guanílico (GMP) mediante modificaciones químicas del anillo de purina. La síntesis de nucleótidos de pirimidina comienza con la formación de ácido orotidílico, cuya descarboxilación da ácido uridílico (UMP). A partir de este último se forma UTP, cuya acilación conduce a la formación de CTP.
Los desoxirribonucleótidos se forman como resultado de la reducción de los ribonucleótidos correspondientes al nivel de difosfatos o trifosfatos. La síntesis de un nucleótido específico de ADN, el ácido timidílico, ocurre por metilación enzimática del ácido desoxiuridílico.
Intercambio iónico. Los compuestos minerales - iones, NH 3 + , K + , Mg 2+ , Fe 2+ , SO 4 2- , PO 4 3- y otras bacterias se obtienen del medio ambiente tanto en estado libre como asociado a otros compuestos orgánicos. sustancias Los cationes y aniones se transportan al interior de la célula bacteriana de diversas formas, descritas en el § 3. La velocidad de penetración de los iones en la célula bacteriana se ve afectada por el pH del medio y la actividad fisiológica de los propios microorganismos.
B) Respiración de las bacterias (metabolismo energético).
Todos los procesos de la vida dependen de la energía, por lo tanto, la obtención de energía es un aspecto extremadamente importante del metabolismo de los procariotas. Obtienen energía de la respiración anaeróbica y aeróbica.
Aliento, o La oxidación biológica es un proceso catabólico de transferencia de electrones de una sustancia donadora a una sustancia aceptora, acompañada de la acumulación de energía en compuestos macroérgicos. . La respiración se lleva a cabo en el proceso de reacciones catabólicas, como resultado de lo cual las sustancias orgánicas complejas, al dividirse, emiten energía y se convierten en compuestos simples. La energía acumulada en sustancias macroérgicas (ATP, GTP, UTP, etc.) se utiliza en reacciones anabólicas.
Según el modo de respiración, los microorganismos se clasifican en aerobios obligados (estrictos), anaerobios obligados y anaerobios facultativos.
aerobios obligados necesitan oxígeno libre. Los compuestos orgánicos (carbohidratos, grasas, proteínas) son donantes de electrones en aerobios quimioorganotróficos patógenos para humanos, y el oxígeno molecular es un aceptor de electrones. El almacenamiento de energía en forma de ATP en aerobios quimioorganotróficos ocurre durante la fosforilación oxidativa de los donantes de electrones. Los aerobios tienen citocromos (participan en la transferencia de electrones), así como enzimas (catalasa, superóxido dismutasa, peroxidasa) que inactivan los radicales de oxígeno tóxicos generados durante la respiración. La superóxido dismutasa inactiva el metabolito más tóxico, el radical superóxido en H 2 O 2 . La enzima catalasa convierte el H 2 O 2 en H 2 O y O 2.
Un grupo especial de aerobios son bacterias microaerófilas, que, aunque necesitan oxígeno para obtener energía, crecen mejor con un mayor contenido de CO 2, por ejemplo, las bacterias del género Campylobacter y Helicobacter.
anaerobios obligados no necesitan oxígeno libre, por el contrario, incluso en pequeñas cantidades, el oxígeno tiene un efecto tóxico sobre ellos. Los donantes de electrones en anaerobios-quimioorganotrofos patógenos humanos son varios compuestos orgánicos (principalmente carbohidratos). El aceptor de electrones en los anaerobios quimioorganotróficos son compuestos orgánicos que contienen oxígeno: ácidos o cetonas, es decir, el aceptor de electrones es el oxígeno asociado con el fragmento orgánico. El almacenamiento de energía en estos procariotas ocurre durante la fosforilación del sustrato. Los anaerobios obligados, por regla general, no tienen citocromos y enzimas que inactivan los radicales de oxígeno (catalasa y superóxido dismutasa negativos).
En los anaerobios quimiolitotróficos que no son patógenos para los humanos, el aceptor de electrones son compuestos inorgánicos que contienen oxígeno: nitratos, sulfatos, carbonatos.
Un grupo especial de anaerobios son las bacterias aerotolerantes que pueden crecer en presencia de oxígeno atmosférico, pero no lo utilizan como aceptor de electrones (por ejemplo, las bacterias del ácido láctico). Los procariotas aerotolerantes a la catalasa y la superóxido dismutasa son positivos.
Anaerobios facultativos capaz de existir tanto en oxígeno como en ambientes anóxicos. Sus donantes de electrones son sustancias orgánicas; Los aceptores de electrones, dependiendo de las condiciones ambientales, son moleculares o se unen al oxígeno en compuestos orgánicos e inorgánicos. Los anaerobios facultativos pueden acumular energía tanto durante la fosforilación oxidativa como del sustrato. Al igual que los aerobios, este grupo de bacterias posee citocromos y enzimas de defensa antioxidantes.
El principal sustrato para la obtención de energía son los carbohidratos, que en procariotas quimioheterotróficas de diferentes tipos de respiración pueden catabolizarse a acetil coenzima A (“ácido acético activado”). Los lípidos y las proteínas pueden actuar como sustratos energéticos, ya que la acetilcoenzima A es también uno de los productos intermedios de su metabolismo (Esquema 2).
El catabolismo de carbohidratos en procariotas quimioorganotróficas incluye: (a) procesos anaeróbicos: glucólisis, la vía de las pentosas fosfato y la vía del cetodesoxifosfogluconato; (b) proceso aeróbico - ciclo del ácido tricarboxílico (ciclo de Krebs). Los procesos anaeróbicos tienen lugar en todos los procariontes, mientras que el proceso aeróbico es característico solo de los aerobios obligados y los anaerobios facultativos. Las vías anaeróbicas se basan en la fosforilación del sustrato, mientras que los procesos aeróbicos se basan en la fosforilación oxidativa.
Definición de conceptos.
La esterilización, la desinfección y los antisépticos son partes integrales de la práctica médica moderna y especialmente quirúrgica. Comprender los principios y la aplicación práctica de estos métodos es esencial porque muchos microorganismos potencialmente patógenos pueden sobrevivir fuera del huésped durante largos períodos de tiempo, exhiben una alta resistencia a los desinfectantes físicos y químicos y se transmiten con relativa facilidad de persona a persona.
Antisépticos- destrucción o prevención del crecimiento de microorganismos patógenos u oportunistas por métodos químicos. Este término suele utilizarse para referirse a la aplicación externa de un preparado químico a tejidos vivos.
antiséptico- una sustancia que inhibe el crecimiento o destruye un microorganismo (sin acción sobre las esporas bacterianas). El término es específico para las sustancias que se utilizan para la acción tópica en los tejidos vivos.
Asepsia significa la ausencia de sepsis, pero en general este término se usa para enfatizar la ausencia de organismos vivos. Métodos asépticos significa cualquier procedimiento diseñado para eliminar organismos vivos y prevenir la recontaminación por ellos. Las modernas técnicas quirúrgicas y microbiológicas se basan en procedimientos asépticos.
biocida- una sustancia que mata todos los microorganismos vivos, tanto patógenos como no patógenos, incluidas las esporas.
Biostato- un agente que previene el crecimiento de microorganismos, pero no necesariamente los mata.
Descontaminación- eliminación de microorganismos sin determinación cuantitativa. Este término es relativo; la eliminación final de microbios se puede lograr mediante esterilización o desinfección.
Desinfección- un proceso que reduce o elimina todos los patógenos excepto las esporas.
Germicida- una sustancia que destruye los microorganismos, especialmente los patógenos. El germicida no destruye las esporas.
sanación- un método por el cual la contaminación microbiana se reduce a un nivel "seguro". Este método se utilizó anteriormente para "purificar" objetos inanimados.
Esterilización- el uso de factores físicos y (o) químicos para la destrucción completa o destrucción de todas las formas de vida microbiana.
Esterilización.
La esterilización se define como la destrucción o eliminación (por filtración) de todos los microorganismos y sus esporas. La esterilización generalmente se lleva a cabo usando calor. La esterilización, siendo una de las rutinas diarias en el trabajo de un laboratorio de microbiología, es un método esencial para garantizar que los cultivos, equipos, utensilios y medios apoyen el crecimiento de solo los microorganismos necesarios, mientras que otros microbios son destruidos. Existen tales tipos de esterilización: calcinación en la llama de un quemador, ebullición, acción con vapor que fluye, vapor a presión en un autoclave, calor seco, pasteurización, tindalización, esterilización química, fría (mecánica).
Elección de los métodos de esterilización.
Al elegir los métodos de esterilización, se deben considerar los siguientes requisitos:
1. Actividad: bactericida, esporicida, tuberculocida, fungicida y virocida.
2. Rapidez del procedimiento: la esterilización debe realizarse lo más rápido posible.
3. Permeabilidad: Los agentes esterilizantes deben poder penetrar a través del embalaje y en el interior del instrumento.
4. Compatibilidad: no debe haber cambios en la estructura o función de los materiales que se esterilizan varias veces.
5. No toxicidad: no debe haber ninguna amenaza para la salud humana y el medio ambiente.
6. Persistencia de material orgánico: la eficacia de la esterilización no debe reducirse en presencia de material orgánico.
7. Adaptabilidad: la capacidad de uso para grandes y pequeños volúmenes de material esterilizado.
8. Control en el tiempo: el ciclo de procesamiento debe controlarse con facilidad y precisión.
9. Precio: costo razonable de equipo, instalación y operación.
 Esterilizadores físicos
Esterilizadores físicos
mojado cálido, que se forma durante el proceso de autoclave a vapor, es el principal agente esterilizante utilizado en los laboratorios de microbiología clínica. Los autoclaves se utilizan para esterilizar medios de cultivo, materiales refractarios y tratar desechos infecciosos. Un esterilizador de vapor, o autoclave, es una cámara presurizada aislada que utiliza vapor saturado para generar altas temperaturas (Figura 1). El aire se elimina de la cámara por desplazamiento de masa o vacío. Los autoclaves más utilizados con sustitución por peso. Se introduce vapor más ligero en la cámara para desplazar el aire más pesado. Una breve exposición al vapor presurizado puede destruir las esporas bacterianas. Para la esterilización rutinaria de medios de cultivo y otros materiales, el tiempo de exposición es de 15 minutos a 121 °C y una presión de 1,5 kg por centímetro cuadrado. Para desechos infecciosos, el tiempo de exposición se aumenta a 30-60 minutos. Además del tiempo y la temperatura adecuados, el contacto directo con el vapor es muy importante para la esterilización. Al manipular material infeccioso, debe asegurarse la máxima penetración de vapor en los desechos. Dicho material debe procesarse a una temperatura de 132ºС. Los antineoplásicos, los productos químicos tóxicos y los radioisótopos que pueden no descomponerse y los productos químicos inestables no deben esterilizarse en autoclave porque pueden evaporarse y propagarse a través de la cámara cuando se exponen al calor.
 Esterilización por calor seco se utiliza para materiales que no pueden esterilizarse con vapor debido a la posibilidad de daño o debido a la impermeabilidad del material al vapor. El calor seco es menos efectivo que el calor húmedo y requiere tiempos de exposición más largos y temperaturas más altas. La esterilización por calor seco se suele realizar en una cabina de calor seco (Fig. 2). El mecanismo de esterilización por calor seco es un proceso oxidativo. Ejemplos de materiales para los que se utiliza la esterilización por calor seco son aceites, polvos, instrumentos afilados y cristalería. El calor seco o la inactivación-esterilización térmica se utilizan como métodos alternativos para el tratamiento de residuos infecciosos.
Esterilización por calor seco se utiliza para materiales que no pueden esterilizarse con vapor debido a la posibilidad de daño o debido a la impermeabilidad del material al vapor. El calor seco es menos efectivo que el calor húmedo y requiere tiempos de exposición más largos y temperaturas más altas. La esterilización por calor seco se suele realizar en una cabina de calor seco (Fig. 2). El mecanismo de esterilización por calor seco es un proceso oxidativo. Ejemplos de materiales para los que se utiliza la esterilización por calor seco son aceites, polvos, instrumentos afilados y cristalería. El calor seco o la inactivación-esterilización térmica se utilizan como métodos alternativos para el tratamiento de residuos infecciosos.
La pasteurización destruye los patógenos calentando rápidamente la sustancia a 71,1 °C durante 15 segundos, seguido de un enfriamiento rápido. La pasteurización no es esterilización porque no todos los microorganismos son sensibles a ella. Este método eliminó la transmisión alimentaria de enfermedades como la tuberculosis gastrointestinal y la fiebre Q.
tindalización es un método de esterilización por calor intermitente que se puede usar para matar todas las bacterias en las soluciones. Debido a que las bacterias en crecimiento se matan fácilmente con hervidos cortos (5 veces en 1 hora durante 5 minutos), todo lo que se necesita hacer es dejar reposar la solución durante un cierto período de tiempo antes de que el calor interrumpa la maduración de las esporas con un pérdida significativa de su resistencia al calor.
Filtración es un proceso que se utiliza para eliminar microbios y partículas microscópicas de soluciones, aire y otros gases. El uso más común de la esterilización por filtración en el laboratorio es para procesar diagnósticos, medios de cultivo, medios de cultivo de tejidos, sueros, soluciones que contienen componentes de suero. Otra aplicación común de la filtración es la esterilización de aire y gases. Los filtros de membrana de plástico o papel, que se distinguen por el diámetro de los poros (de aproximadamente 12 a 0,22 µm) y se utilizan para la separación mecánica, también sirven para recolectar microbios de líquidos para examen microscópico o cultivo directamente en el filtro cuando se colocan sobre una superficie empapada de nutrientes. medio.
irradiación ultravioleta es un tipo de radiación de ondas electromagnéticas que actúa sobre el ácido nucleico celular. Los microorganismos son muy sensibles a la acción de los rayos ultravioleta con una longitud de onda de 254 nm. La luz ultravioleta es la más utilizada para matar microorganismos en el aire o en las superficies. Otros usos son la esterilización en frío de ciertos productos químicos y plásticos con fines farmacéuticos, la esterilización de suero para cultivos celulares y la desinfección del agua. Una desventaja significativa de la radiación ultravioleta como esterilizador es su incapacidad para penetrar materiales.
radiación ionizante en el espectro electromagnético, tiene un efecto letal sobre los microorganismos. Este espectro incluye microondas, rayos γ, rayos X y flujo de electrones. El efecto letal de la radiación ionizante ocurre debido a la acción directa sobre la molécula objetivo, como resultado de lo cual se transfiere energía a la molécula; y debido a la acción indirecta - la difusión de radicales.
energía ultrasónica con baja frecuencia inactiva los microorganismos en soluciones acuosas. El efecto físico de la sonicación se debe a la cavitación. Los limpiadores ultrasónicos y otros dispositivos a menudo se usan para limpiar instrumentos, pero no se consideran esterilizadores. Sin embargo, la combinación de ultrasonido con tratamiento químico mata los microorganismos.
esterilizadores químicos
2 % glutaraldehído Como esterilizador químico líquido, anteriormente se usaba ampliamente para procesar material médico y quirúrgico que no se puede esterilizar por calor o irradiación. El glutaraldehído también se utiliza en la preparación de vacunas.
Desinfección.
La desinfección se puede realizar por métodos químicos o por ebullición. La ebullición es un método eficaz para desinfectar instrumentos como agujas y jeringas si no se dispone de un autoclave. Los instrumentos médicos previamente limpiados deben hervirse durante 20 minutos. La desinfección química se utiliza para equipos sensibles al calor que pueden dañarse con las altas temperaturas. Los desinfectantes químicos como los componentes de cloro, el alcohol etílico e isopropílico, los componentes de amonio cuaternario y el glutaraldehído son ampliamente utilizados.
Desinfectantes químicos.
Alcohol (etílico e isopropílico), disuelto en agua a una concentración de 60-85%, es muy eficaz en la desinfección. Los alcoholes son bactericidas, fungicidas y tuberculocidas, pero no afectan a las esporas. El alcohol etílico tiene un espectro más amplio de actividad virocida que el alcohol isopropílico, por lo que es más efectivo contra los virus lipofílicos e hidrofílicos.
Solución 37% formaldehído, que se llama formalina, se puede usar como esterilizante, mientras que sus concentraciones de 3-8% se pueden usar como desinfectantes.
Fenol en su forma pura no se utiliza como desinfectante debido a su toxicidad, la capacidad de inducir el desarrollo de tumores y la corrosión. Los derivados del fenol, en los que un grupo funcional (cloro, bromo, alquilo, bencilo, fenilo, amilo) reemplaza uno de los átomos de hidrógeno en el anillo aromático, se usan ampliamente como desinfectantes. Esta sustitución reduce las desventajas del fenol. Los componentes del fenol matan a los microbios debido a la inactivación de los sistemas enzimáticos, la precipitación de las proteínas y la alteración de la pared y la membrana celular. Por lo general, se utilizan concentraciones de 2-5%, las concentraciones más bajas requieren una exposición más prolongada.
halógenos. Solo se utilizan cloro y yodo para la desinfección en la práctica de laboratorio. Debido al hecho de que el cloro es un poderoso agente oxidante, se cree que mata los microbios por oxidación. Se cree que el yodo mata los microorganismos al reaccionar con los grupos N-H y S-H de los aminoácidos, así como con el grupo fenólico del aminoácido tirosina y los dobles enlaces carbono-carbono de los ácidos grasos insaturados. El tratamiento convencional consiste en pulverizar una solución de formaldehído al 2-5% en presencia de vapor a una temperatura de 60-80ºC.
Antisépticos.
Los antisépticos se pueden encontrar en laboratorios microbiológicos, principalmente en sustancias que se utilizan para lavarse las manos. En los casos en que el personal médico brinda atención de emergencia a los pacientes que utilizan sustancias que contienen agentes antibacterianos, esto reduce la cantidad de infecciones hospitalarias. Los productos químicos más comunes que se encuentran en los lavados de manos son alcoholes, gluconato de clorhexidina, yodóforos, cloroxilenol y triclosán.
Los métodos tradicionales de tratamiento de desechos y basuras son la incineración y la esterilización por vapor.
Incendio es el método de elección para el manejo de desechos y escombros. Este método hace que los desechos no sean infecciosos y también cambia su forma y tamaño. La esterilización es un método eficaz de tratamiento de residuos, pero no cambia su forma. La esterilización con vapor en un autoclave a 121 °C durante un mínimo de 15 minutos mata todas las formas de vida microbiana, incluido un gran número de esporas bacterianas. Este tipo de esterilización completa también se puede realizar mediante calor seco a 160-170ºC durante 2-4 horas. Sin embargo, se debe asegurar que el calor seco esté en contacto con el material a esterilizar. Por lo tanto, las botellas que contienen líquido deben sellarse sin apretar con corchos o hisopos de algodón para que el vapor y el calor puedan intercambiarse con el aire de las botellas. Los contenedores de riesgo biológico que contengan desechos deben estar bien amarrados. El material de riesgo biológico esterilizado debe sellarse en recipientes debidamente etiquetados.
Esterilización por vapor (autoclave). La basura infecciosa se considera descontaminada cuando el número de bacterias vegetativas, hongos, micobacterias y virus que contienen lípidos disminuye en 6 lg veces, y las endosporas bacterianas en 4 lg veces.
Nutrición de las bacterias.
Alimento. Bajo la nutrición de una célula bacteriana, se debe entender el proceso de absorción y asimilación de materia plástica y energía como resultado de reacciones transformadoras. . Los tipos de nutrición de los procariotas son complejos y variados. Difieren según la forma en que los nutrientes ingresan a la célula bacteriana, las fuentes de carbono y nitrógeno, la forma en que se obtiene la energía y la naturaleza de los donantes de electrones.
Transporte de nutrientes al interior de la célula. puede llevarse a cabo por 3 mecanismos: difusión pasiva, difusión facilitada y transporte activo.
Difusión pasiva es un proceso no específico dependiente de la energía que se lleva a cabo a lo largo del gradiente de concentración de las sustancias (una sustancia de un ambiente con una concentración más alta ingresa pasivamente, de acuerdo con las leyes de la ósmosis, a un ambiente con una concentración más baja). Por difusión pasiva, una cantidad limitada de sustancias ingresan a la célula bacteriana, algunos iones, monosacáridos. La tasa de transferencia de sustancias durante la difusión pasiva es insignificante y depende de la lipofilia y el tamaño de las moléculas transportadas.
Difusión facilitada es un transporte no volátil de sustancias a lo largo de un gradiente de concentración con la ayuda de enzimas permeasas. Las permeasas son proteínas de membrana específicas que facilitan el paso de sustancias a través de la membrana citoplasmática. La permeasa fija sobre sí misma una molécula de la sustancia transferida, junto con la cual supera la membrana citoplasmática, después de lo cual se disocia el complejo "sustancia-permeasa". La permeasa liberada se utiliza para conducir otras moléculas. En procariotas, solo el glicerol ingresa a la célula por difusión facilitada. En este caso, la concentración intracelular de glicerol corresponde a la del exterior de la célula. La difusión facilitada es más característica de los microorganismos eucariotas.
transporte activo- esta es una transferencia de sustancias dependiente de la energía hacia la célula contra un gradiente de concentración con la ayuda de enzimas específicas. La gran mayoría de las sustancias (iones, carbohidratos, aminoácidos, lípidos, etc.) ingresan a la célula bacteriana por transporte activo. El transporte activo puede tener lugar: (1) sin modificación química de la sustancia transportada; (2) con modificación química.
Las bacterias son microorganismos procarióticos de estructura celular. Sus tamaños son de 0,1 a 30 micras. Los microbios son extremadamente comunes. Viven en el suelo, el aire, el agua, la nieve e incluso en las fuentes termales, en el cuerpo de los animales, así como en el interior de los organismos vivos, incluido el cuerpo humano.
La distribución de bacterias en especies se basa en varios criterios, entre los cuales se tienen en cuenta con mayor frecuencia la forma de los microorganismos y su distribución espacial. Entonces, según la forma de las células, las bacterias se dividen en:
Coci - micro-, diplo-, estrepto-, estafilococos, así como sarcinas;
En forma de bastón: monobacterias, diplobacterias y estreptobacterias;
Especies enrevesadas - vibrios y espiroquetas.
El determinante de Bergey sistematiza todas las bacterias conocidas de acuerdo con los principios de identificación de bacterias que han encontrado la más amplia distribución en la bacteriología práctica, basándose en las diferencias en la estructura de la pared celular y en relación con la tinción de Gram. La descripción de las bacterias se da por grupos (secciones), que incluyen familias, géneros y especies; en algunos casos, los grupos incluyen clases y órdenes. Las bacterias patógenas para los humanos se incluyen en un pequeño número de grupos.
La clave distingue cuatro categorías principales de bacterias:
Gracillicutes [del lat. gracilis, agraciado, delgado, + cutis, piel] - especie con una pared celular delgada, teñida gram negativo;
firmicutes [del lat. flrmus, fuerte, + cutis, piel] - bacterias con una pared celular gruesa, tinción Gram positivas;
Tenericutes [del lat. tener, suave, + cutis, piel] - bacterias que carecen de una pared celular(micoplasmas y otros miembros de la clase Mollicutes)
Mendosicutes [del lat. mendosus, irregular, + cutis, piel] - arqueobacterias (reductoras de metano y sulfato, halófilas, termófilas y arqueobacterias, desprovistas de pared celular).
Grupo 2 Determinante de Burgey. Bacterias gramnegativas aerobias y microaerófilas móviles contorneadas y curvadas. Las especies patógenas para humanos se incluyen en los géneros Campylobacter, Helicobacters Spirillum.
Grupo 3 del determinante de Bergey. Bacterias gramnegativas no móviles (raramente móviles). No contiene especies patógenas.
Grupo 4 del determinante de Burgey. Cocos y bacilos aerobios y microaerófilos gramnegativos. Las especies patógenas para humanos se incluyen en las familias Legionellaceae, Neisseriaceae y Pseudomonada-ceae, el grupo también incluye bacterias patógenas y oportunistas de los géneros Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Francisella, Kingella y Moraxella.
Grupo 5 del determinante de Bergey. Bacilos gramnegativos anaerobios facultativos. El grupo está formado por tres familias - Enterobacteriaceae, Vibrionaceae y Pasteurellaceae, cada una de las cuales incluye especies patógenas, así como bacterias patógenas y oportunistas de los géneros Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella y Streptobacillus.
Grupo 6 del determinante de Bergey. Bacterias anaerobias gramnegativas rectas, curvas y espirales. Las especies patógenas y oportunistas se incluyen en los géneros Bacteroides, Fusobacterium, Porphoromonas y Prevotelta.
Grupo 7 del determinante de Bergey. Bacterias que realizan reducción de la disimilación de sulfato o azufre No incluye especies patógenas.
Grupo 8 del determinante de Bergey. Cocos Gram negativos anaerobios. Incluye bacterias oportunistas del género Veillonella.
Grupo 9 del determinante de Bergey. Rickettsia y clamidia. Tres familias: Rickettsiaceae, Bartonellaceae y Chlamydiaceae, cada una de las cuales contiene especies patógenas para los humanos.
Los grupos 10 y 11 de la guía de Burgey incluyen bacterias fototróficas anoxi y oxigénicas que no son patógenas para los humanos.
Grupo 12 del determinante de Burgey. Bacterias aerobias quimiolitotrofas y organismos relacionados. Combina bacterias oxidantes y nitrificantes de azufre-hierro y manganeso que no causan daño a los humanos.
Los grupos 13 y 14 de la guía de Burgey incluyen bacterias en gemación y/o crecimiento y bacterias formadoras de vaina. Representado por especies de vida libre, no patógenas para los humanos;
Los grupos 15 y 16 de la guía de Burgey unen bacterias deslizantes que no forman cuerpos fructíferos y los forman. Los grupos no incluyen especies patógenas para los humanos.
Grupo 17 del determinante de Burgey. cocos grampositivos. Incluye especies oportunistas de los géneros Enterococcus Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus.
Grupo 18 del determinante de Burgey. Cocos y bacilos grampositivos formadores de esporas. Incluye bacilos patógenos condicionalmente patógenos de los géneros Clostridium y Bacillus.
Grupo 19 del determinante de Burgey. Bastoncillos grampositivos formadores de esporas de forma regular. Incluyendo especies oportunistas de los géneros Erysipelothrix y Listeria.
Grupo 20 del determinante de Burgey. Bastoncillos grampositivos formadores de esporas de forma irregular. El grupo incluye especies patógenas y oportunistas de los géneros Actinomyces, Corynebacterium Gardnerella, Mobiluncus, etc.
Grupo 21 del determinante de Burgey. Micobacterias. Incluye el único género Mycobacterium, que combina especies patógenas y oportunistas.
Grupos 22-29. Actinomicetos. Entre numerosas especies, solo los actinomicetos nocardioformes (Grupo 22) de los géneros Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi y Terrabacter son capaces de causar lesiones en humanos.
Grupo 30 del determinante de Burgey. Micoplasmas. Las especies incluidas en los géneros Acholeplasma, Mycoplasma y Ureaplasma son patógenas para los humanos.
Los grupos restantes del determinante de Bergey - bacterias metanogénicas (31), bacterias sulfato reductoras (32 arqueobacterias aeróbicas extremadamente halófilas (33), arqueobacterias sin pared celular (34), termófilas e hipertermófilas extremas, que metabolizan azufre (35) - no contienen especies patógenas para los humanos.
Las bacterias son el grupo de organismos más antiguo que existe actualmente en la Tierra. Las primeras bacterias probablemente aparecieron hace más de 3.500 millones de años y durante casi mil millones de años fueron los únicos seres vivos de nuestro planeta. Dado que estos fueron los primeros representantes de la vida silvestre, su cuerpo tenía una estructura primitiva.
Con el tiempo, su estructura se volvió más compleja, pero incluso hoy en día las bacterias se consideran los organismos unicelulares más primitivos. Curiosamente, algunas bacterias aún conservan las características primitivas de sus antiguos ancestros. Esto se observa en bacterias que viven en manantiales calientes de azufre y limos anóxicos en el fondo de los embalses.
La mayoría de las bacterias son incoloras. Sólo unos pocos son de color púrpura o verde. Pero las colonias de muchas bacterias tienen un color brillante, que se debe a la liberación de una sustancia coloreada al medio ambiente oa la pigmentación de las células.
El descubridor del mundo de las bacterias fue Anthony Leeuwenhoek, un naturalista holandés del siglo XVII, quien creó por primera vez un microscopio de lupa perfecto que aumenta los objetos entre 160 y 270 veces.
Las bacterias se clasifican como procariotas y se separan en un reino separado: las bacterias.
forma del cuerpo
Las bacterias son organismos numerosos y diversos. Se diferencian en la forma.
| nombre de la bacteria | Forma de bacterias | Imagen de bacterias |
| cocos | esférico | |
| Bacilo |  | en forma de varilla |
| Vibrión | coma curva | |
| espirilum |  | Espiral |
| estreptococos |  | Cadena de cocos |
| estafilococos |  | Racimos de cocos |
| diplococos | Dos bacterias redondas encerradas en una cápsula viscosa. |
formas de transporte
Entre las bacterias hay formas móviles e inmóviles. Los móviles se mueven mediante contracciones ondulatorias o con la ayuda de flagelos (hilos helicoidales retorcidos), que consisten en una proteína flagelina especial. Puede haber uno o más flagelos. Están ubicados en algunas bacterias en un extremo de la célula, en otras, en dos o en toda la superficie.

Pero el movimiento también es inherente a muchas otras bacterias que no tienen flagelos. Entonces, las bacterias cubiertas con moco en el exterior son capaces de moverse por deslizamiento.
Algunas bacterias del agua y del suelo sin flagelos tienen vacuolas de gas en el citoplasma. Puede haber 40-60 vacuolas en una celda. Cada uno de ellos está lleno de gas (presumiblemente nitrógeno). Al regular la cantidad de gas en las vacuolas, las bacterias acuáticas pueden hundirse en la columna de agua o subir a su superficie, mientras que las bacterias del suelo pueden moverse en los capilares del suelo.
Hábitat
Debido a la simplicidad de la organización y la falta de pretensiones, las bacterias están ampliamente distribuidas en la naturaleza. Las bacterias se encuentran en todas partes: incluso en una gota del agua de manantial más pura, en los granos de tierra, en el aire, en las rocas, en las nieves polares, en las arenas del desierto, en el fondo del océano, en el aceite extraído de grandes profundidades e incluso en agua caliente. agua de manantial con una temperatura de unos 80ºС. Viven en plantas, frutas, en varios animales y en humanos en los intestinos, la boca, las extremidades y en la superficie del cuerpo.
Las bacterias son los seres vivos más pequeños y numerosos. Debido a su pequeño tamaño, penetran fácilmente en cualquier grieta, hendidura y poro. Muy resistente y adaptado a diversas condiciones de existencia. Toleran el secado, el frío extremo, el calentamiento hasta 90ºС, sin perder viabilidad.
Prácticamente no hay lugar en la Tierra donde no se encuentren bacterias, pero en diferentes cantidades. Las condiciones de vida de las bacterias son variadas. Algunos de ellos necesitan oxígeno del aire, otros no lo necesitan y pueden vivir en un ambiente libre de oxígeno.
En el aire: las bacterias ascienden a la atmósfera superior hasta 30 km. y más.
Especialmente muchos de ellos en el suelo. Un gramo de suelo puede contener cientos de millones de bacterias.
En agua: en las capas superficiales de agua de embalses abiertos. Las bacterias acuáticas beneficiosas mineralizan los residuos orgánicos.
En los organismos vivos: las bacterias patógenas ingresan al cuerpo desde el ambiente externo, pero solo en condiciones favorables causan enfermedades. Simbiótico vive en los órganos digestivos, ayudando a descomponer y asimilar los alimentos, sintetizar vitaminas.
Estructura externa
La célula bacteriana está vestida con una capa densa especial: la pared celular, que realiza funciones protectoras y de apoyo, y también le da a la bacteria una forma característica permanente. La pared celular de una bacteria se asemeja a la cáscara de una célula vegetal. Es permeable: a través de él, los nutrientes pasan libremente a la célula y los productos metabólicos salen al medio ambiente. Las bacterias a menudo desarrollan una capa protectora adicional de moco, una cápsula, sobre la pared celular. El grosor de la cápsula puede ser muchas veces mayor que el diámetro de la propia célula, pero puede ser muy pequeño. La cápsula no es una parte obligatoria de la célula, se forma según las condiciones en las que ingresan las bacterias. Evita que las bacterias se sequen.
En la superficie de algunas bacterias hay flagelos largos (uno, dos o muchos) o vellosidades cortas y delgadas. La longitud de los flagelos puede ser muchas veces mayor que el tamaño del cuerpo de la bacteria. Las bacterias se mueven con la ayuda de flagelos y vellosidades.

Estructura interna
Dentro de la célula bacteriana hay un citoplasma inmóvil denso. Tiene una estructura en capas, no hay vacuolas, por lo que varias proteínas (enzimas) y nutrientes de reserva se encuentran en la sustancia misma del citoplasma. Las células bacterianas no tienen núcleo. En la parte central de sus células se concentra una sustancia portadora de información hereditaria. Bacterias, - ácido nucleico - ADN. Pero esta sustancia no está enmarcada en el núcleo.
La organización interna de una célula bacteriana es compleja y tiene sus propias características específicas. El citoplasma está separado de la pared celular por la membrana citoplasmática. En el citoplasma, se distinguen la sustancia principal, o matriz, los ribosomas y un pequeño número de estructuras de membrana que realizan una variedad de funciones (análogos de las mitocondrias, el retículo endoplásmico, el aparato de Golgi). El citoplasma de las células bacterianas a menudo contiene gránulos de varias formas y tamaños. Los gránulos pueden estar compuestos por compuestos que sirven como fuente de energía y carbono. Las gotas de grasa también se encuentran en la célula bacteriana.
En la parte central de la célula se localiza la sustancia nuclear, el ADN, que no está separada del citoplasma por una membrana. Este es un análogo del núcleo: el nucleoide. El nucleoide no tiene membrana, nucléolo y un conjunto de cromosomas.
metodos de nutricion
Las bacterias tienen diferentes formas de alimentarse. Entre ellos se encuentran los autótrofos y los heterótrofos. Los autótrofos son organismos que pueden formar sustancias orgánicas de forma independiente para su nutrición.
Las plantas necesitan nitrógeno, pero ellas mismas no pueden absorber el nitrógeno del aire. Algunas bacterias combinan moléculas de nitrógeno en el aire con otras moléculas, dando como resultado sustancias disponibles para las plantas.
Estas bacterias se asientan en las células de las raíces jóvenes, lo que conduce a la formación de engrosamientos en las raíces, llamados nódulos. Dichos nódulos se forman en las raíces de las plantas de la familia de las leguminosas y algunas otras plantas.

Las raíces proporcionan carbohidratos a las bacterias, y las bacterias les dan a las raíces sustancias que contienen nitrógeno que pueden ser absorbidas por la planta. Su relación es mutuamente beneficiosa.
Las raíces de las plantas secretan muchas sustancias orgánicas (azúcares, aminoácidos y otras) de las que se alimentan las bacterias. Por lo tanto, especialmente muchas bacterias se asientan en la capa del suelo que rodea las raíces. Estas bacterias convierten los residuos de plantas muertas en sustancias disponibles para la planta. Esta capa de suelo se llama rizosfera.
Hay varias hipótesis sobre la penetración de las bacterias del nódulo en los tejidos de la raíz:
- a través del daño al tejido epidérmico y cortical;
- a través de los pelos de la raíz;
- sólo a través de la membrana celular joven;
- debido a bacterias compañeras que producen enzimas pectinolíticas;
- debido a la estimulación de la síntesis de ácido B-indolacético a partir del triptófano, que siempre está presente en las secreciones de las raíces de las plantas.
El proceso de introducción de bacterias del nódulo en el tejido radicular consta de dos fases:
- infección de los pelos de la raíz;
- proceso de formación de nódulos.
En la mayoría de los casos, la célula invasora se multiplica activamente, forma los llamados hilos de infección y ya en forma de tales hilos se mueve hacia los tejidos de la planta. Las bacterias del nódulo que han surgido del hilo de infección continúan multiplicándose en el tejido huésped.

Llenas de células de bacterias de nódulos que se multiplican rápidamente, las células vegetales comienzan a dividirse intensamente. La conexión de un nódulo joven con la raíz de una planta leguminosa se lleva a cabo gracias a haces vasculares y fibrosos. Durante el período de funcionamiento, los nódulos suelen ser densos. En el momento de la manifestación de la actividad óptima, los nódulos adquieren un color rosado (debido al pigmento de legoglobina). Solo aquellas bacterias que contienen legoglobina son capaces de fijar nitrógeno.
Las bacterias del nódulo crean decenas y cientos de kilogramos de fertilizantes nitrogenados por hectárea de suelo.
Metabolismo
Las bacterias difieren entre sí en el metabolismo. Para algunos, va con la participación de oxígeno, para otros, sin su participación.
La mayoría de las bacterias se alimentan de sustancias orgánicas preparadas. Solo algunos de ellos (azul-verde o cianobacterias) son capaces de crear sustancias orgánicas a partir de sustancias inorgánicas. Desempeñaron un papel importante en la acumulación de oxígeno en la atmósfera terrestre.
Las bacterias absorben sustancias del exterior, desgarran sus moléculas, ensamblan su caparazón a partir de estas partes y reponen su contenido (así es como crecen) y desechan las moléculas innecesarias. El caparazón y la membrana de la bacteria le permiten absorber solo las sustancias adecuadas.

Si la cubierta y la membrana de la bacteria fueran completamente impermeables, ninguna sustancia entraría en la célula. Si fueran permeables a todas las sustancias, el contenido de la célula se mezclaría con el medio, la solución en la que vive la bacteria. Para la supervivencia de las bacterias se necesita un caparazón que permita el paso de las sustancias necesarias, pero no de las que no se necesitan.
La bacteria absorbe los nutrientes que se encuentran cerca de ella. ¿Qué pasa después? Si puede moverse de forma independiente (moviendo el flagelo o empujando el moco hacia atrás), entonces se mueve hasta que encuentra las sustancias necesarias.

Si no puede moverse, espera hasta que la difusión (la capacidad de las moléculas de una sustancia para penetrar en el espesor de las moléculas de otra sustancia) le traiga las moléculas necesarias.
Las bacterias, junto con otros grupos de microorganismos, realizan un enorme trabajo químico. Mediante la transformación de diversos compuestos, reciben la energía y los nutrientes necesarios para su actividad vital. Los procesos metabólicos, las formas de obtención de energía y la necesidad de materiales para construir las sustancias de su organismo en las bacterias son diversos.
Otras bacterias satisfacen todas las necesidades de carbono necesarias para la síntesis de sustancias orgánicas del cuerpo a expensas de compuestos inorgánicos. Se llaman autótrofos. Las bacterias autótrofas son capaces de sintetizar sustancias orgánicas a partir de inorgánicas. Entre ellos se distinguen:
quimiosíntesis
El uso de energía radiante es la forma más importante, pero no la única, de crear materia orgánica a partir de dióxido de carbono y agua. Se sabe que las bacterias no utilizan la luz solar como fuente de energía para dicha síntesis, sino la energía de los enlaces químicos que se producen en las células de los organismos durante la oxidación de ciertos compuestos inorgánicos: sulfuro de hidrógeno, azufre, amoníaco, hidrógeno, ácido nítrico, compuestos ferrosos de hierro y manganeso. Usan la materia orgánica formada usando esta energía química para construir las células de su cuerpo. Por lo tanto, este proceso se llama quimiosíntesis.
El grupo más importante de microorganismos quimiosintéticos son las bacterias nitrificantes. Estas bacterias viven en el suelo y llevan a cabo la oxidación del amoníaco, formado durante la descomposición de los residuos orgánicos, a ácido nítrico. Este último, reacciona con los compuestos minerales del suelo, se convierte en sales de ácido nítrico. Este proceso se desarrolla en dos fases.
Las bacterias del hierro convierten el hierro ferroso en óxido. El hidróxido de hierro formado se asienta y forma el llamado mineral de hierro de pantano.
Algunos microorganismos existen debido a la oxidación del hidrógeno molecular, proporcionando así una forma autótrofa de nutrición.
Un rasgo característico de las bacterias del hidrógeno es la capacidad de cambiar a un estilo de vida heterótrofo cuando se les proporcionan compuestos orgánicos y en ausencia de hidrógeno.
Por lo tanto, los quimioautótrofos son autótrofos típicos, ya que sintetizan de forma independiente los compuestos orgánicos necesarios a partir de sustancias inorgánicas y no los toman preparados de otros organismos, como los heterótrofos. Las bacterias quimioautotróficas difieren de las plantas fototróficas en su total independencia de la luz como fuente de energía.
fotosíntesis bacteriana
Algunas bacterias de azufre que contienen pigmentos (púrpura, verde), que contienen pigmentos específicos, las bacterioclorofilas, pueden absorber la energía solar, con la ayuda de la cual el sulfuro de hidrógeno se divide en sus organismos y proporciona átomos de hidrógeno para restaurar los compuestos correspondientes. Este proceso tiene mucho en común con la fotosíntesis y difiere solo en que en las bacterias moradas y verdes, el sulfuro de hidrógeno (ocasionalmente ácidos carboxílicos) es un donante de hidrógeno, y en las plantas verdes es agua. En esos y otros, el desdoblamiento y transferencia de hidrógeno se realiza debido a la energía de los rayos solares absorbidos.
Tal fotosíntesis bacteriana, que ocurre sin la liberación de oxígeno, se llama fotorreducción. La fotorreducción de dióxido de carbono está asociada con la transferencia de hidrógeno no del agua, sino del sulfuro de hidrógeno:
6CO 2 + 12H 2 S + hv → C6H 12 O 6 + 12S \u003d 6H 2 O
La importancia biológica de la quimiosíntesis y la fotosíntesis bacteriana a escala planetaria es relativamente pequeña. Solo las bacterias quimiosintéticas juegan un papel importante en el ciclo del azufre en la naturaleza. Absorbido por las plantas verdes en forma de sales de ácido sulfúrico, el azufre se restaura y pasa a formar parte de las moléculas de proteínas. Además, durante la destrucción de residuos vegetales y animales muertos por bacterias putrefactas, el azufre se libera en forma de sulfuro de hidrógeno, que es oxidado por las bacterias del azufre a azufre libre (o ácido sulfúrico), que forma sulfitos disponibles para las plantas en el suelo. Las bacterias quimio y fotoautótrofas son esenciales en el ciclo del nitrógeno y el azufre.
esporulación
Las esporas se forman dentro de la célula bacteriana. En el proceso de formación de esporas, una célula bacteriana pasa por una serie de procesos bioquímicos. La cantidad de agua libre en él disminuye, la actividad enzimática disminuye. Esto asegura la resistencia de las esporas a condiciones ambientales adversas (alta temperatura, alta concentración de sal, secado, etc.). La formación de esporas es característica de sólo un pequeño grupo de bacterias.

Las esporas no son una etapa esencial en el ciclo de vida de las bacterias. La esporulación comienza solo con la falta de nutrientes o la acumulación de productos metabólicos. Las bacterias en forma de esporas pueden permanecer latentes durante mucho tiempo. Las esporas bacterianas resisten la ebullición prolongada y la congelación muy prolongada. Cuando se dan las condiciones favorables, la disputa germina y se hace viable. Las esporas bacterianas son adaptaciones para sobrevivir en condiciones adversas.
reproducción
Las bacterias se reproducen dividiendo una célula en dos. Habiendo alcanzado cierto tamaño, la bacteria se divide en dos bacterias idénticas. Luego, cada uno de ellos comienza a alimentarse, crece, se divide, etc.

Después de la elongación de la célula, se forma gradualmente un tabique transversal y luego las células hijas divergen; en muchas bacterias, bajo ciertas condiciones, las células después de la división permanecen conectadas en grupos característicos. En este caso, dependiendo de la dirección del plano de división y del número de divisiones, surgen diferentes formas. La reproducción por gemación ocurre en las bacterias como una excepción.
Bajo condiciones favorables, la división celular en muchas bacterias ocurre cada 20-30 minutos. Con una reproducción tan rápida, la descendencia de una bacteria en 5 días puede formar una masa que puede llenar todos los mares y océanos. Un simple cálculo muestra que se pueden formar 72 generaciones (720.000.000.000.000.000.000 células) por día. Si se traduce en peso - 4720 toneladas. Sin embargo, esto no sucede en la naturaleza, ya que la mayoría de las bacterias mueren rápidamente bajo la influencia de la luz solar, el secado, la falta de alimentos, el calentamiento a 65-100ºС, como resultado de la lucha entre especies, etc.
La bacteria (1), habiendo absorbido suficiente alimento, aumenta de tamaño (2) y comienza a prepararse para la reproducción (división celular). Su ADN (en una bacteria la molécula de ADN está cerrada en un anillo) se duplica (la bacteria produce una copia de esta molécula). Ambas moléculas de ADN (3.4) parecen estar adheridas a la pared bacteriana y, cuando se alargan, las bacterias divergen hacia los lados (5.6). Primero se divide el nucleótido, luego el citoplasma.
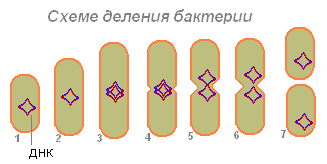
Después de la divergencia de dos moléculas de ADN en las bacterias, aparece una constricción que divide gradualmente el cuerpo de la bacteria en dos partes, cada una de las cuales contiene una molécula de ADN (7).
Ocurre (en el bacilo del heno), dos bacterias se unen y se forma un puente entre ellas (1,2).

El ADN se transporta de una bacteria a otra a través de un puente (3). Una vez en una bacteria, las moléculas de ADN se entrelazan, se unen en algunos lugares (4), después de lo cual intercambian secciones (5).
El papel de las bacterias en la naturaleza.
Circulación
Las bacterias son el eslabón más importante en la circulación general de sustancias en la naturaleza. Las plantas crean sustancias orgánicas complejas a partir de dióxido de carbono, agua y sales minerales del suelo. Estas sustancias regresan al suelo con hongos muertos, plantas y cadáveres de animales. Las bacterias descomponen sustancias complejas en simples, que son reutilizadas por las plantas.
Las bacterias destruyen la materia orgánica compleja de plantas muertas y cadáveres de animales, excreciones de organismos vivos y diversos desechos. Al alimentarse de estas sustancias orgánicas, las bacterias saprofitas de la descomposición las convierten en humus. Estos son el tipo de camilleros de nuestro planeta. Así, las bacterias participan activamente en el ciclo de las sustancias en la naturaleza.
formación del suelo
Dado que las bacterias se distribuyen en casi todas partes y se encuentran en grandes cantidades, determinan en gran medida los diversos procesos que ocurren en la naturaleza. En otoño, las hojas de los árboles y arbustos caen, los brotes de hierba que se encuentran sobre el suelo mueren, las ramas viejas se caen y, de vez en cuando, caen los troncos de los árboles viejos. Todo esto se convierte gradualmente en humus. En 1 cm 3. La capa superficial del suelo forestal contiene cientos de millones de bacterias saprofitas del suelo de varias especies. Estas bacterias convierten el humus en varios minerales que las raíces de las plantas pueden absorber del suelo.
Algunas bacterias del suelo pueden absorber nitrógeno del aire y utilizarlo en los procesos vitales. Estas bacterias fijadoras de nitrógeno viven solas o se instalan en las raíces de las leguminosas. Habiendo penetrado en las raíces de las leguminosas, estas bacterias provocan el crecimiento de las células de las raíces y la formación de nódulos en ellas.
Estas bacterias liberan compuestos de nitrógeno que utilizan las plantas. Las bacterias obtienen carbohidratos y sales minerales de las plantas. Así, existe una estrecha relación entre la leguminosa y la bacteria del nódulo, lo cual es útil tanto para uno como para el otro organismo. Este fenómeno se llama simbiosis.
Gracias a su simbiosis con las bacterias del nódulo, las leguminosas enriquecen el suelo con nitrógeno, lo que ayuda a aumentar los rendimientos.
Distribución en la naturaleza
Los microorganismos son ubicuos. Las únicas excepciones son los cráteres de volcanes activos y pequeñas áreas en los epicentros de bombas atómicas detonadas. Ni las bajas temperaturas de la Antártida, ni los chorros hirvientes de los géiseres, ni las soluciones salinas saturadas en las piscinas de sal, ni la fuerte insolación de los picos de las montañas, ni la dura radiación de los reactores nucleares interfieren con la existencia y el desarrollo de la microflora. Todos los seres vivos interactúan constantemente con los microorganismos, siendo muchas veces no solo sus almacenes, sino también distribuidores. Los microorganismos son los nativos de nuestro planeta, desarrollando activamente los sustratos naturales más increíbles.
microflora del suelo
La cantidad de bacterias en el suelo es extremadamente grande: cientos de millones y miles de millones de individuos en 1 gramo. Son mucho más abundantes en el suelo que en el agua y el aire. El número total de bacterias en los suelos varía. El número de bacterias depende del tipo de suelo, su condición, la profundidad de las capas.
En la superficie de las partículas del suelo, los microorganismos se encuentran en pequeñas microcolonias (20-100 células cada una). A menudo se desarrollan en el espesor de coágulos de materia orgánica, en raíces de plantas vivas y moribundas, en capilares delgados y en el interior de bultos.
La microflora del suelo es muy diversa. Aquí se encuentran diferentes grupos fisiológicos de bacterias: bacterias putrefactivas, nitrificantes, fijadoras de nitrógeno, sulfurosas, etc. entre ellas hay aerobias y anaerobias, formas de esporas y no esporas. La microflora es uno de los factores de formación del suelo.
El área de desarrollo de microorganismos en el suelo es la zona adyacente a las raíces de las plantas vivas. Se llama rizosfera, y la totalidad de los microorganismos contenidos en ella se llama microflora de la rizosfera.
Microflora de reservorios
El agua es un entorno natural donde los microorganismos crecen en grandes cantidades. La mayoría de ellos ingresan al agua desde el suelo. Un factor que determina la cantidad de bacterias en el agua, la presencia de nutrientes en ella. Las más limpias son las aguas de pozos artesianos y manantiales. Los embalses abiertos y los ríos son muy ricos en bacterias. El mayor número de bacterias se encuentra en las capas superficiales del agua, más cerca de la costa. A medida que aumenta la distancia de la costa y aumenta la profundidad, el número de bacterias disminuye.
El agua pura contiene de 100 a 200 bacterias por 1 ml, mientras que el agua contaminada contiene de 100 a 300 mil o más. Hay muchas bacterias en el sedimento del fondo, especialmente en la capa superficial, donde las bacterias forman una película. Hay una gran cantidad de bacterias de azufre y hierro en esta película, que oxidan el sulfuro de hidrógeno a ácido sulfúrico y, por lo tanto, evitan que los peces mueran. Hay más formas portadoras de esporas en el limo, mientras que las formas no portadoras de esporas predominan en el agua.
En términos de composición de especies, la microflora del agua es similar a la microflora del suelo, pero también se encuentran formas específicas. Destruyendo diversos desechos que han caído al agua, los microorganismos van realizando poco a poco la llamada depuración biológica del agua.
Microflora del aire
La microflora del aire es menos numerosa que la microflora del suelo y del agua. Las bacterias se elevan en el aire con el polvo, pueden permanecer allí por un tiempo y luego se asientan en la superficie de la tierra y mueren por falta de nutrición o bajo la influencia de los rayos ultravioleta. La cantidad de microorganismos en el aire depende del área geográfica, el terreno, la estación, la contaminación por polvo, etc. Cada mota de polvo es portadora de microorganismos. La mayoría de las bacterias en el aire sobre las empresas industriales. El aire en el campo es más limpio. El aire más limpio está sobre bosques, montañas, espacios nevados. Las capas superiores del aire contienen menos gérmenes. En la microflora del aire hay muchas bacterias pigmentadas y portadoras de esporas que son más resistentes que otras a los rayos ultravioleta.
Microflora del cuerpo humano.
El cuerpo de una persona, incluso uno completamente sano, es siempre portador de microflora. Cuando el cuerpo humano entra en contacto con el aire y el suelo, una variedad de microorganismos, incluidos patógenos (bacilos del tétanos, gangrena gaseosa, etc.), se depositan en la ropa y la piel. Las partes expuestas del cuerpo humano son las más frecuentemente contaminadas. E. coli, los estafilococos se encuentran en las manos. Hay más de 100 tipos de microbios en la cavidad oral. La boca, con su temperatura, humedad, residuos de nutrientes, es un ambiente excelente para el desarrollo de microorganismos.
El estómago tiene una reacción ácida, por lo que la mayor parte de los microorganismos mueren. A partir del intestino delgado, la reacción se vuelve alcalina, es decir, favorable para los microbios. La microflora en el intestino grueso es muy diversa. Cada adulto excreta alrededor de 18 mil millones de bacterias diariamente con sus excrementos, es decir, más individuos que personas en el mundo.
Los órganos internos que no están conectados con el entorno externo (cerebro, corazón, hígado, vejiga, etc.) generalmente están libres de microbios. Los microbios ingresan a estos órganos solo durante la enfermedad.
Bacterias en el ciclismo
Los microorganismos en general y las bacterias en particular juegan un papel importante en los ciclos biológicamente importantes de la materia en la Tierra, llevando a cabo transformaciones químicas que son completamente inaccesibles para las plantas o los animales. Varias etapas del ciclo de los elementos son llevadas a cabo por organismos de diferentes tipos. La existencia de cada grupo separado de organismos depende de la transformación química de los elementos llevada a cabo por otros grupos.
ciclo del nitrógeno
La transformación cíclica de los compuestos nitrogenados juega un papel primordial en el suministro de las formas necesarias de nitrógeno a varios organismos de la biosfera en términos de necesidades nutricionales. Más del 90% de la fijación total de nitrógeno se debe a la actividad metabólica de ciertas bacterias.
El ciclo del carbono
La transformación biológica del carbono orgánico en dióxido de carbono, acompañada de la reducción del oxígeno molecular, requiere la actividad metabólica conjunta de varios microorganismos. Muchas bacterias aerobias llevan a cabo la oxidación completa de las sustancias orgánicas. En condiciones aeróbicas, los compuestos orgánicos se descomponen inicialmente por fermentación y los productos orgánicos finales de la fermentación se oxidan aún más por la respiración anaeróbica si están presentes aceptores de hidrógeno inorgánicos (nitrato, sulfato o CO2).
ciclo del azufre
Para los organismos vivos, el azufre está disponible principalmente en forma de sulfatos solubles o compuestos orgánicos reducidos de azufre.
El ciclo del hierro
Algunos depósitos de agua dulce contienen altas concentraciones de sales de hierro reducidas. En tales lugares, se desarrolla una microflora bacteriana específica: bacterias de hierro, que oxidan el hierro reducido. Participan en la formación de minerales de hierro de pantano y fuentes de agua ricas en sales de hierro.
Las bacterias son los organismos más antiguos, apareciendo hace unos 3.500 millones de años en el Arcaico. Durante unos 2500 millones de años, dominaron la Tierra, formaron la biosfera y participaron en la formación de una atmósfera de oxígeno.
Las bacterias son uno de los organismos vivos más simples (a excepción de los virus). Se cree que son los primeros organismos que aparecieron en la Tierra.
bacterias- Estos son organismos unicelulares, libres de clorofila, que se reproducen vegetativamente por división, con menos frecuencia por enlace, a veces formando esporas intracelulares.
El tamaño de las bacterias se mide en micras y, con raras excepciones, oscila entre 0,06-0,3 y 3-5 μ. Una gota de agua puede albergar fácilmente varios cientos de millones de bacterias.
La forma de una célula bacteriana es bastante uniforme. Se conocen tres formas principales de bacterias: redondas, en forma de varilla y enrevesadas con numerosas e imperceptibles transiciones entre ellas. Anton De Bari los comparó en sentido figurado con una bola de billar, un lápiz y un sacacorchos.
Las bacterias que tienen forma redonda se llaman cocos. Difieren en tamaño y posición relativa. Los cocos conectados en pares se llaman diplococos, pero los conectados en forma de collar se llaman estreptococos. Al dividirse, alternando en dos direcciones mutuamente perpendiculares, se forman tetracocos. Si la división es correcta y se repite en tres direcciones perpendiculares entre sí, las conexiones celulares se forman en forma de paquetes: estas son las llamadas sardinas. Dividiéndose en diferentes direcciones sin mucha regularidad, los cocos forman racimos desordenados que se asemejan a un cepillo de uvas. Se llaman estafilococos.
Las bacterias con forma de bastón son algo más diversas en su apariencia. Pueden ser con extremos truncados o redondeados, cilíndricos, en forma de barril o de limón y, por así decirlo, con una constricción en el medio, a menudo elipsoidal, que difieren solo en su ancho y largo. Los palos pueden ser rectos, curvos, simples, unidos en pares o en cadena, cortos o fuertemente alargados. Las bacterias con forma de bastoncillo, en las que la longitud es dos veces o más que el ancho, se denominan bacilos; si la diferencia entre el largo y el ancho es pequeña, entonces se llaman bacterias.
Las bacterias con forma de rizo difieren no solo en longitud y grosor, sino también en el número y la naturaleza de los rizos. Las bacterias ligeramente curvadas (el rizo no supera 1/4 de vuelta de la espiral) se denominan vibrio, las bacterias con uno o más rizos regulares grandes se denominan espirilla. Las bacterias largas y delgadas de forma retorcida con numerosos rizos pequeños, a veces con grandes curvaturas de todo el filamento, se denominan espiroquetas.
La estructura de las bacterias.
Por la simplicidad de su organización y tamaño insignificante, las bacterias pertenecen a los seres más elementales y se encuentran en los peldaños más bajos de la vida. A pesar de los tremendos avances en ciencia y tecnología, aún no se han resuelto todas las preguntas sobre la estructura de las bacterias.
El cuerpo de las bacterias consta de una coraza y un protoplasma de contenido ético, impregnado de savia celular. El caparazón de las bacterias es delgado, incoloro, su estructura no se distingue bajo un microscopio. Para verlo, recurren a métodos artificiales de procesamiento. La vaina subyace a la forma exterior de la célula y parece ser una conocida defensa contra condiciones adversas. Envolviendo libremente el contenido de la célula, gracias a su elasticidad, permite la libre circulación de bacterias, a menudo acompañada de movimientos enérgicos de todo el cuerpo.
Las capas exteriores de la concha, que absorben agua, a menudo se hinchan y forman una masa pegajosa gelatinosa, que alcanza un tamaño notable. A medida que las capas exteriores se vuelven mucilaginosas, la cubierta se renueva continuamente a expensas del protoplasma. La cubierta adhesiva enfriada se llama cápsula. La intensidad de la formación de mocos y cápsulas depende de las características de la alimentación y en ocasiones puede ser muy importante. La cápsula mucosa se forma no solo cerca de cada celda por separado, sino también en muchas celdas conectadas en una colonia y encerradas, por así decirlo, en una cápsula común. Estas colonias viscosas de bacterias se llaman zoogles. La formación de cápsulas no es característica de todos los tipos de bacterias.
movimiento de bacterias
La capacidad de movimiento espontáneo es inherente solo a algunos grupos de bacterias. Las bacterias se mueven por medio de flagelos o cilios. Los flagelos parecen filamentos más o menos largos. Son muy delicados, delgados, se rompen con facilidad y no son visibles al microscopio sin una tinción especial. Su diámetro no supera 1/20 del diámetro del cuerpo bacteriano.
Las formas móviles de bacterias se dividen en los siguientes grupos:
- monotrichous - solo hay un flagelo polar,
- lophotrichous - hay un haz de flagelos en un extremo de la celda,
- los flagelos peritricos se encuentran en toda la superficie del cuerpo.
La naturaleza de la ubicación de los flagelos en el cuerpo de la bacteria también determina la naturaleza de su movimiento: rectilíneo o aleatorio. La motilidad de las bacterias depende de una serie de condiciones: temperatura, composición del medio nutritivo, productos de su actividad vital, etc. Las formas móviles de bacterias no están equipadas con flagelos en todas las etapas de su desarrollo y no en todas las condiciones de crecimiento.
esporulación
En el cuerpo de muchas bacterias, en ciertos períodos de su desarrollo, aparecen formaciones redondas o elipsoidales: soportes. Suelen completar el ciclo de desarrollo de las bacterias. El tamaño de las esporas en comparación con el tamaño de las células que las produjeron puede variar ampliamente.
Los soportes no se forman en todos los tipos de bacterias. Están rodeados por un caparazón bien aislado, casi impermeable al agua y son las formaciones más estables entre todo el mundo viviente. Por lo tanto, a menudo resisten la ebullición durante varias horas y la exposición prolongada al vapor seco a temperaturas de 120 a 140 °. Las esporas de muchos bacilos conservan su viabilidad después de una larga estancia a una temperatura de -190° e incluso a una temperatura de hidrógeno líquido (-253°). También son resistentes a la acción de los productos químicos: venenos. Todo esto hace que sea extremadamente difícil combatir las especies de bacterias patógenas de esporas.
Una espora madura puede permanecer viable durante décadas. Por lo general, la germinación de esporas ocurre después de un cierto período de latencia y está asociada con la influencia de condiciones externas. Todo el proceso de esporulación ocurre en un día o menos. Después de que la espora madura, la célula que la produjo muere gradualmente y sale la espora madura. Durante la germinación, se hincha, se enriquece en agua y sale una plántula, vestida con una cáscara delgada.
Reproducción de bacterias
Habiendo alcanzado el estado de madurez y límite de crecimiento, las bacterias comienzan a multiplicarse por simple división. Durante la división, aparece un tabique en la parte media del cuerpo de la bacteria, que luego se divide y separa dos nuevas células. La disposición secuencial de las particiones durante la división es diferente para diferentes bacterias. En formas en forma de varilla, se ubica perpendicular al eje largo, en formas esféricas, las particiones se pueden ubicar en uno, dos o tres planos mutuamente perpendiculares, razón por la cual se forman formas como estreptococos, tetracocos y sarcinas. .
La tasa de reproducción de las bacterias depende de una serie de condiciones y puede ser muy diferente. Cuanto más favorables son las condiciones externas para la existencia de bacterias, más rápido se produce su división. En condiciones normales, el número de bacterias se duplica aproximadamente cada media hora. Si siempre sucediera sin obstáculos, entonces la cantidad de bacterias de una célula alcanzaría proporciones colosales. Según el microbiólogo Kohn, la descendencia de un bacilo en cinco días podría llenar todos los mares y océanos. Sin embargo, esto nunca ha sucedido y nunca sucederá. El ciclo de vida de las bacterias está limitado por ciertas condiciones externas, más allá de las cuales la reproducción se ralentiza o se detiene por completo. La falta de nutrición, los productos metabólicos nocivos, la competencia de varias especies, etc., tienen un efecto perjudicial sobre las bacterias. En condiciones adversas, mueren en masa.
Clasificación de las bacterias
La posición de las bacterias en el sistema de los seres vivos aún no está bien definida. En general, se acepta que las bacterias son parte del mundo vegetal y que los hongos y las algas son los organismos más cercanos a ellas. Las características morfológicas de las bacterias en la mayoría de los casos se limitan a algunas formas: esféricas, cilíndricas, espirales. La extraordinaria sencillez y la elementalidad de su organización externa dificultan su clasificación. Es imposible determinar la especie de una bacteria basándose únicamente en caracteres morfológicos. La sistemática científica se basa en la morfología y la historia del desarrollo, pero para clasificar las bacterias es necesario conocer no solo la morfología, sino también sus características fisiológicas y bioquímicas. En este sentido, se establecen los siguientes: la proporción de bacterias a oxígeno, las condiciones de temperatura, la formación de pigmento, la licuefacción de la gelatina, la formación de ácidos y gas en los azúcares, el cambio en la leche con el crecimiento de bacterias en ella, la formación de indol, sulfuro de hidrógeno, amoníaco, la reducción de nitratos a nitritos o nitrógeno libre. Sin embargo, esto no siempre es suficiente para determinar el tipo de bacteria.
Existen varios sistemas de clasificación para las bacterias, pero todos son condicionales y se alejan más o menos de la clasificación natural. La consideración de estos sistemas o al menos uno de ellos en este caso no es necesaria incluso cuando se aplica a bacterias fitopatógenas. Solo cabe decir que en la actualidad casi todas las bacterias fitopatógenas se agrupan en los géneros Pseudomonas, Xanthomonas, Bacterium y Erwinia.
Recientemente, M. V. Gorlenko (1961) propuso el siguiente sistema de clasificación para las bacterias fitopatógenas de la clase Eubacteriales:
YO. Familia Mycobacteriaceae(Chester, 1901) - bacterias no móviles (sin flagelos):
- 1er género - Gorynebacterium (Leman y Neumann, 1896) - (bacterias grampositivas;
- 2do género Aplanobacterium (Smith, 1905, Geshich, 1956) - bacterias gramnegativas.
II. Familia Pseudomonadaceae(Wilson et al., 1917) - bacterias con flagelos (polares):
- 1er género - Pseudomonas (Migula, 1900) - bacterias sin teñir y fluorescentes;
- 2do género - Xanthomonas (Dawson, 1839) - bacterias con colonias coloreadas.
tercero Familia Bacteriaceas(Kon, 1872) - bacterias móviles con flagelos peritricos que no forman soportes:
- 1er género - Bacterium (Ehrenberger, 1828) - formas no teñidas que no forman pectinasas y protopectinasas;
- 2do género - Pectobacterium (Waldy, 1945) - formas no teñidas que forman pectinasa y protopectinasa;
- 3er género - Chromobacterium (Bergonzini, 1881) - formas coloreadas.
IV. Familia Bacillaceae(Fischer, 1895) - bacterias móviles, bacilos formadores de esporas:
- 1er género - Bacillus (Kon, 1832) - las células no se hinchan o se hinchan ligeramente durante la formación de esporas;
- 2do género - Clostridium (Praznovsky, 1880) - las células se hinchan durante la formación de esporas.
En el sistema anterior, se omite el género Erwinia generalmente aceptado hasta ahora. De él se aísla un género especial Pectobacterium, que incluye bacterias con flagelos peritricos y actividad pectolítica. Aquellas de las bacterias fitopatógenas que no poseen esta capacidad se asignan al género Bacterium. Este sistema, racional en sí mismo, es nuevo y aún no ha entrado en la vida cotidiana, por lo que en este trabajo nos adherimos a la clasificación en la que se le da su lugar al género Erwinia. Este nombre genérico de bacteria fitopatógena es ampliamente utilizado en la literatura especializada tanto en nuestro país como en el exterior.
Es imposible determinar el tipo de bacteria sin el uso de medios nutrientes artificiales. Al respecto, se puede señalar que al cultivar bacterias, éstas forman colonias muy características. En este caso, se puede usar una apariencia para juzgar las especies de bacterias.
Si encuentra un error, resalte un fragmento de texto y haga clic en Ctrl+Intro.
1.5. Clasificación moderna de bacterias En la taxonomía moderna de bacterias, se ha desarrollado una situación que también es característica de la clasificación de otros organismos: se ha avanzado en la creación de un sistema de clasificación filogenético que refleja las principales direcciones del desarrollo evolutivo y la relación de representantes de ciertos taxones, pero los fenómenos artificiales conservan su significado.- clasificaciones típicas, más convenientes para la identificación de microorganismos. En la actualidad, no existe ningún sistema evolutivo detallado de los procariotas y, muy probablemente, la solución de este problema no es cuestión de un futuro cercano. Las características de los procariotas en el campo de la organización morfológica, fisiológica, bioquímica y genética indican la inaplicabilidad de los principios bien desarrollados utilizados en la construcción de un sistema de organismos superiores para ellos. Sin detenernos en los aspectos históricos del problema de la taxonomía bacteriana, cabe señalar que el sistema de clasificación filogenética más aceptable para procariotas es un sistema basado en la coincidencia de la secuencia de nucleótidos en el ARNr 16S. Este sistema formó la base de la segunda edición de la enciclopedia de procariotas de varios volúmenes - Bergey's Manual of Systematic Bacteriology (Bergey's Guide to Systematic Bacteria), cuyo primer volumen se publicó en 2001. En este trabajo, todos los procariotas se dividen en 26 "ramas" filogenéticas (grupos) basadas en la estructura de su 16S-rRNA; 23 "ramas" están representadas por eubacterias y tres por arqueobacterias. Cabe destacar que un gran número de estos grupos filogenéticos contienen especies procarióticas que no han sido aisladas como cultivos puros y por lo tanto aún no han sido estudiadas en detalle. Para los representantes de estas especies, actualmente solo se conocen las secuencias de nucleótidos del ARNr 16S. De los 23 grupos de eubacterias, dos grupos filogenéticos están representados por bacterias grampositivas, los grupos restantes son gramnegativos. Las bacterias Gram negativas consisten en un gran grupo de Proteobacterias (Proteobacteria) y 20 grupos de otras bacterias con este tipo de pared celular. En la Tabla 1 se proporciona una breve descripción de las proteobacterias, a las que las mitocondrias y los cloroplastos de la mayoría de los eucariotas están más cerca en la composición de 16S-rRNA. 2. Las proteobacterias son un grupo de bacterias gramnegativas muy heterogéneo en términos morfológicos, fisiológicos y bioquímicos. Los representantes de este grupo se caracterizan por todo tipo de metabolismo energético y nutrición. Las células de la mayoría de las especies de Proteobacteria tienen forma de bastón, esférica o vibrioide; se reproducen principalmente por fisión binaria, pero algunas especies se caracterizan por brotar y formar cuerpos fructíferos en un ciclo celular complejo. Este grupo contiene bacterias móviles debido a flagelos e inmóviles. Con respecto al oxígeno molecular, las proteobacterias se clasifican en aerobios obligados, anaerobios obligados y facultativos. El grupo Proteobacteria se divide en cinco subgrupos según las diferencias en el ARNr 16S: alfa, beta, gamma, delta y épsilon. Además de las proteobacterias, las eubacterias gramnegativas incluyen los siguientes grupos principales de eubacterias: hidrogenotermófilos, bacterias filamentosas verdes, bacterias verdes del azufre, cianobacterias, espiroquetas, citofagos, bacteroides, clamidias, planctomicetos, deinococos, cloroflexos, fusobacterias, fibrobacterias, termodesulfobacterias, etc. Grupos filogenéticos bacterias grampositivas - Actinobacteria y Firmicutes. El grupo Actinobacteria (“rama actinomycete”) está representado por los siguientes géneros de bacterias con un alto contenido de pares GC en el ADN: Geodermatophilus, Frankia, Streptomyces, Arthrobacter, Micrococcus, Actinomyces, Bifidobacterium, Propionibacterium, Actinoplanes, Nocardia, Rhodococcus, Corynebacterium , micobacterias. El grupo Firmicutes ("rama clostridial" - principalmente bacterias Gram-positivas con un bajo contenido de pares GC en el ADN) consta de los siguientes géneros: Clostridium, Lactococcus, Pediococcus, Streptococcus, Enterococcus, Leuconostoc, Listeria, Caryophanon, Staphylococcus, Sarcina , Sporosarcina, Bacillus, Desulfotomaculum, Heliobacterium, Mycoplasma, Ureaplasma, etc. Las arqueobacterias se dividen en tres grupos filogenéticos: Crenarchaeota, Euryarchaeota y Korarchaeota. El grupo Crenarchaeota consiste en bacterias extremadamente termófilas, la mayoría de cuyos representantes metabolizan el azufre, algunos reducen los iones de hierro y molibdeno. El grupo Euryarchaeota incluye arqueobacterias metanogénicas anaerobias obligadas, así como termófilas y halófilas extremas. El grupo Korarchaeota está formado por arqueobacterias que viven en manantiales calientes de azufre. Hasta la fecha, ninguno de los representantes de este grupo (que poseen un rRNA 16S similar) ha sido aislado como cultivo puro, por lo que sus características fenotípicas no han sido suficientemente estudiadas. Concluyendo la consideración de las ramas filogenéticas de los procariotas, cabe señalar que el sistema filogenético propuesto, basado en el estudio de las secuencias de nucleótidos de un solo gen de ARN ribosomal, no es más que uno de los sistemas técnicamente convenientes y desarrollados para ordenar numerosos organismos con el fin de identificarlos, por lo que es lógico construir una taxonomía correcta de bacterias teniendo en cuenta que esta característica no es posible. La clasificación fenotípica de bacterias más reconocida y utilizada es la que se presenta en la novena edición de Burgey's Guide to Bacteria. En esta edición, las bacterias se dividen en cuatro categorías principales (divisiones) según la estructura de la capa límite celular: 1) Gracilicutes (del latín cutes - piel, gracilis - delgado) - eubacterias gramnegativas con paredes celulares; 2) Firmicutes (del lat. firmus - fuerte) - eubacterias grampositivas con paredes celulares; 3) Tenericutes (del latín tener - suave, tierno) - eubacteria, sin paredes celulares; 4) Mendosicutes (del latín mendosus - erróneo) - arqueobacterias, cuyas paredes celulares difieren de estructuras similares de otros procariotas. El departamento de Gracilicutes incluye bacterias de varias morfologías con una pared celular Gram-negativa. La reproducción se produce principalmente por fisión binaria, algunas bacterias se reproducen por gemación. No se forma endospora. La mayoría son móviles: hay todo tipo de movimiento de bacterias, con la ayuda de flagelos, deslizamiento, flexión. La división incluye bacterias aeróbicas, anaeróbicas y anaeróbicas facultativas; bacterias fotótrofas y quimiotróficas. El departamento se divide en tres clases: Scotobacteria, Oxyphotobacteria, Anoxyphotobacteria. La clase Scotobacteria incluye bacterias gramnegativas que no utilizan la energía de la luz con fines metabólicos, sino que la reciben solo como resultado de reacciones redox. El nombre de la clase proviene del griego. sсotos - oscuridad. Esta es la clase más grande de bacterias. La clase Anoxifotobacteria incluye bacterias moradas, bacterias verdes y heliobacterias, que realizan la fotosíntesis anoxigénica (sin liberación de oxígeno molecular). La clase Oxyphotobacteria está representada por cianobacterias y proclorofitas que realizan la fotosíntesis oxigénica (con liberación de oxígeno molecular). Este tipo de fotosíntesis es similar a la fotosíntesis que ocurre en las plantas. El departamento de Firmicutes incluye bacterias con una pared celular Gram-positiva. Las células pueden tener diferentes formas: bastones, cocos, filamentosos, ramificados. Algunos representantes forman endosporas. La mayoría de ellos están inmóviles; las formas móviles tienen flagelos peritricos. El departamento incluye bacterias aeróbicas, anaeróbicas y anaeróbicas facultativas. El departamento consta de dos clases: Firmibacteria, Thallobacteria. La clase Firmibacteria incluye una gran cantidad de bacterias Gram-positivas "no ramificadas". La clase Thallobacteria incluye bacterias cuyas células son capaces de "ramificarse". El departamento de Tenericutes está representado por bacterias que no tienen pared celular. Debido a la ausencia de una pared celular, la forma de las células no es constante: en un cultivo puro de una especie, las células cocoides, en forma de varilla, filamentosas, en forma de pera, en forma de disco y otras están presentes simultáneamente. La reproducción de las bacterias incluidas en este apartado se produce por fisión binaria, gemación. La tinción de Gram es negativa. Es característica la formación de pequeñas colonias que crecen en agar. Pueden ser saprofitos, parásitos o patógenos. El departamento consta de una clase Mollicutes (micoplasmas). La división Mendosicutes está formada por bacterias con una pared celular rígida pero que carecen de peptidoglicano de mureína. La mayoría de los representantes son anaerobios estrictos, muchos de los cuales tienen flagelos. Las especies se caracterizan por la diversidad ecológica y metabólica, la capacidad de vivir en condiciones extremas. El departamento consta de una clase: Archaebacteria. Como parte de cuatro departamentos (categorías principales), se han identificado 35 grupos (o secciones) de bacterias, que se caracterizarán en mayor o menor medida en capítulos posteriores. Los siguientes grupos pertenecen al departamento Gracilicutes. Grupo 1. Espiroquetas. Grupo 2. Bacterias gramnegativas aerobias (o microaerófilas), móviles, espirales (o vibrioides). Grupo 3. Bacterias curvas Gram negativas inmóviles o raramente móviles. Grupo 4. Cocos y bacilos aerobios (o microaerófilos) gramnegativos. Grupo 5. Bastoncillos aerobios gramnegativos opcionales. Grupo 6. Gram-negativos anaerobios bacilos rectos, curvos o espirales. Grupo 7. Bacterias que realizan reducción de la disimilación de azufre o sulfato. Grupo 8. Cocos anaerobios gramnegativos. Grupo 9. Rickettsia y clamidia. Grupo 10. Bacterias fototróficas anoxigénicas. Grupo 11. Bacterias fototróficas oxigénicas. Grupo 12. Bacterias aerobias quimiolitotróficas y organismos relacionados. Grupo 13. Bacterias en gemación y (o) formadoras de crecimiento. Grupo 14. Bacterias con vainas. Grupo 15. Bacterias deslizantes no fotosintéticas que no forman cuerpos fructíferos. Grupo 16. Bacterias deslizantes que forman cuerpos fructíferos. El departamento Firmicutes incluye: Grupo 17. Cocos grampositivos. Grupo 18. Bacilos y cocos grampositivos que forman endosporas. Grupo 19. Bastoncillos grampositivos de forma correcta, que no forman esporas. Grupo 20. Bastoncillos grampositivos de forma irregular que no forman esporas. Grupo 21. Micobacterias. Grupos 22–29. Actinomicetos. Al departamento Tenericutes pertenece: Grupo 30. Micoplasmas. División Mendosicutes incluye: Grupo 31. Metanógenos. Grupo 32. Arqueobacterias sulfatoreductoras. Grupo 33. Arqueobacterias extremadamente halófilas (halobacterias). Grupo 34. Arqueobacterias sin pared celular. Grupo 35. Arqueobacterias metabolizadoras de azufre extremadamente termófilas e hipertermófilas. En conclusión, se debe enfatizar que la mayoría de los microorganismos existentes en las comunidades naturales todavía necesitan ser aislados en cultivos puros. Se cree que en la actualidad solo se puede cultivar el 0,1% de la diversidad microbiana total, y no es posible cultivar e identificar a los restantes representantes de bacterias, aunque ya se han aislado y descrito en cultivo puro unas 5 mil especies de procariotas.