Միկրոօրգանիզմների դասակարգումն ըստ դրանց հիմնական ձևերի. Բակտերիաները, դրանց բազմազանությունը. Կառուցվածք. Կենսունակություն. Հիմնական տերմիններ՝ բակտերիաներ
Բակտերիաների սնուցում.
Սնուցում.
պասիվ դիֆուզիա
Հեշտացված դիֆուզիոն
ակտիվ տրանսպորտ
Առաջին դեպքում սննդանյութի մոլեկուլը կազմում է համալիր պերիպլազմային տիեզերական սպիտակուցի հետ, որը փոխազդում է կոնկրետ ցիտոպլազմային մեմբրանի պերմեազի հետ։ Ցիտոպլազմային մեմբրանի միջով էներգիայից կախված ներթափանցումից հետո «սուբստրատ-պերիպլազմ պրոտեին-պերմեազ» համալիրը տարանջատվում է սուբստրատի մոլեկուլի արտազատման հետ:
Փոխադրվող նյութի քիմիական փոփոխությամբ ակտիվ փոխադրման ժամանակ իրադարձությունների շղթան ներառում է. 2) ֆոսֆորիլացված ֆերմենտ-2 սուբստրատի մոլեկուլների միջոցով ցիտոպլազմային մեմբրանի մակերեսին կապելը. (3) ենթաշերտի մոլեկուլի էներգիայից կախված տեղափոխումը ցիտոպլազմա. (4) ֆոսֆատային խմբի տեղափոխում սուբստրատի մոլեկուլին. (5) «սուբստրատ-ֆերմենտ» համալիրի տարանջատումը ցիտոպլազմայում. Ֆոսֆորիլացման պատճառով սուբստրատի մոլեկուլները կուտակվում են բջիջների ցիտոպլազմայում և չեն կարողանում դուրս գալ դրանցից։
Բակտերիաների դասակարգումը ըստ սննդի տեսակների.
Սննդանյութերի ընդունման եղանակովբակտերիաները դասակարգվում են հոլոֆիտներև Հոլոզոյան. Հոլոֆիտ բակտերիաներ (հունարենից. հոլոներ- ամբողջական և ֆիտիկոս- կապված բույսերի հետ) չեն կարողանում շրջակա միջավայր արտանետել սուբստրատները քայքայող ֆերմենտները, ինչի արդյունքում նրանք սննդանյութերը սպառում են բացառապես լուծված, մոլեկուլային տեսքով: Հոլոզոյան բակտերիաներ (հունարենից. հոլոներ- ամբողջական և զոյկոս- կապված կենդանիների հետ), ընդհակառակը, նրանք ունեն էկզոֆերմենտների համալիր, որոնք ապահովում են արտաքին սնուցում - սուբստրատների քայքայումը բակտերիալ բջիջից դուրս գտնվող մոլեկուլներին: Դրանից հետո սննդանյութերի մոլեկուլները մտնում են հոլոզոյան բակտերիաների ներս։
Ածխածնի աղբյուրովմեկուսացված բակտերիայից ավտոտրոֆներև հետերոտրոֆներ. Ավտոտրոֆներ (հունարենից. ավտոմեքենաներ- ինքս ինձ, ավար- սննդամթերք) ածխածնի երկօքսիդը (CO 2) օգտագործվում է որպես ածխածնի աղբյուր, որից սինթեզվում են ածխածին պարունակող բոլոր նյութերը։ Համար հետերոտրոֆներ (հունարեն գետերոսից - մեկ այլ և ավար- սնունդ) ածխածնի աղբյուրները մոլեկուլային ձևով տարբեր օրգանական նյութեր են (ածխաջրեր, պոլիհիդրային սպիրտներ, ամինաթթուներ, ճարպաթթուներ): Հետերոտրոֆիայի ամենաբարձր աստիճանը բնորոշ է պրոկարիոտներին, որոնք կարող են ապրել միայն այլ կենդանի բջիջների ներսում (օրինակ՝ ռիկեցիա և քլամիդիա):
Ազոտի աղբյուրովպրոկարիոտները բաժանվում են 3 խմբի. 2) բակտերիաներ, որոնք օգտագործում են անօրգանական ազոտ ամոնիումի աղերից, նիտրիտներից կամ նիտրատներից. 3) բակտերիաներ, որոնք յուրացնում են օրգանական միացություններում պարունակվող ազոտը (ամինաթթուներ, պուրիններ, պիրիմիդիններ և այլն).
Ըստ էներգիայի աղբյուրիբակտերիաները բաժանվում են ֆոտոտրոֆներև քիմոտրոֆներ. ֆոտոտրոֆ բակտերիաներ Ինչպես բույսերը, կարողանում են օգտագործել արեգակնային էներգիան։ Ֆոտոտրոֆ պրոկարիոտները մարդկանց մոտ հիվանդություն չեն առաջացնում։ Քիմիոտրոֆ բակտերիաներ էներգիա ստանալ ռեդոքս ռեակցիաներում:
Էլեկտրոնների դոնորների բնույթով լիտոտրոֆներ(հունարենից. լիտոս- քար) և օրգանոտրոֆներ. ժամը լիտոտրոֆներ (քիմոլիտոտրոֆներ ) անօրգանական նյութերը հանդես են գալիս որպես էլեկտրոնների դոնորներ (H 2, H 2 S, NH 3, ծծումբ, CO, Fe 2+ և այլն): Էլեկտրոնների դոնորներ օրգանոտրոֆներ (քիմոօրգանոտրոֆներ ) օրգանական միացություններ են՝ ածխաջրեր, ամինաթթուներ և այլն։
Մարդկանց համար պաթոգեն բակտերիաների մեծ մասն ունի քիմոօրգանոտրոֆիկ (քիմիեթերոտրոֆ) տեսակի սնուցում. քիմոլիտոտրոֆիկ (քիմիաավտոտրոֆիկ) տեսակը ավելի քիչ տարածված է:
Օրգանական միացություններ սինթեզելու ունակությամբքիմոտրոֆ բակտերիաները դասակարգվում են նախատրոֆներ, ավքսոտրոֆներև հիպոտրոֆներ. պրոտոտրոֆ բակտերիաներ սինթեզել բոլոր անհրաժեշտ օրգանական նյութերը գլյուկոզայի և ամոնիումի աղերից: Բակտերիաները կոչվում են աուկսոտրոֆներ եթե նրանք չեն կարողանում նշված միացություններից որևէ օրգանական նյութ սինթեզել. Նյութափոխանակության ակտիվության կորստի ծայրահեղ աստիճանը կոչվում է թերսնուցում: հիպոտրոֆիկ բակտերիաներ ապահովում են իրենց կենսական գործունեությունը` վերակազմավորելով հյուրընկալողի կառուցվածքները կամ մետաբոլիտները:
Բացի ածխածնից և ազոտից, բակտերիաների լիարժեք կյանքի համար անհրաժեշտ են ծծումբ, ֆոսֆոր և մետաղական իոններ: Ծծմբի աղբյուրներն են ամինաթթուները (ցիստեին, մեթիոնին), վիտամինները, կոֆակտորները (բիոտին, լիպոաթթու և այլն), սուլֆատները։ Ֆոսֆորի աղբյուրներն են նուկլեինաթթուները, ֆոսֆոլիպիդները, ֆոսֆատները։ Բավականաչափ բարձր կոնցենտրացիաներում բակտերիաներին անհրաժեշտ է մագնեզիում, կալիում, կալցիում, երկաթ; շատ ավելի փոքր քանակությամբ՝ ցինկ, մանգան, նատրիում, մոլիբդեն, պղինձ, նիկել, կոբալտ:
աճի գործոններ-Սրանք նյութեր են, որոնք բակտերիաները չեն կարող ինքնուրույն սինթեզել, բայց դրանք խիստ կարիք ունեն: Որպես աճի գործոն կարող են հանդես գալ ամինաթթուները, ազոտային հիմքերը, վիտամինները, ճարպաթթուները, երկաթի պորֆիրինները և այլ միացություններ: Բակտերիաների կենսագործունեության համար օպտիմալ պայմաններ ստեղծելու համար սննդարար միջավայրում պետք է ավելացվեն աճի գործոններ:
Նյութափոխանակություն, էներգիայի փոխակերպում
Ա) կառուցողական նյութափոխանակություն.
Բակտերիաների սնուցման պարտադիր փուլը սննդանյութերի յուրացումն է, այսինքն՝ դրանց ընդգրկումը փոփոխված կամ փոփոխված ձևով սինթետիկ ռեակցիաներում՝ բջջային բաղադրիչների և կառուցվածքների վերարտադրության համար:
Սպիտակուցային նյութափոխանակությունբակտերիաների դեպքում այն կարող է ընթանալ 3 փուլով՝ առաջնային սպիտակուցի քայքայում, սպիտակուցի երկրորդային քայքայում և սպիտակուցի սինթեզ: Սպիտակուցի մոլեկուլների առաջնային տարրալուծումը դեպի պեպտոններ իրականացվում է էկզոֆերմենտների՝ էկզոպրոտեազների միջոցով, որոնք թողարկվում են բակտերիաների կողմից շրջակա միջավայր: Երկրորդային քայքայումը տեղի է ունենում էնդոֆերմենտների (էնդոպրոտեազների) ազդեցության ներքո, որոնք ունեն բոլոր բակտերիաները։ Այս գործընթացը տեղի է ունենում բակտերիալ բջջի ներսում և բաղկացած է պեպտիդների տրոհումից մինչև դրանց բաղկացուցիչ ամինաթթուներ: Վերջիններս կարող են օգտագործվել անփոփոխ կամ ենթարկվել քիմիական փոխակերպումների (դեամինացիա, դեկարբոքսիլացում և այլն), որոնց արդյունքում առաջանում են ամոնիակ, ինդոլ, ջրածնի սուլֆիդ, կետոթթուներ, ալկոհոլ, ածխաթթու և այլն: Այդ միացությունների հայտնաբերումը մանրէաբանության մեջ ունի ախտորոշիչ նշանակություն։
Սպիտակուցների տրոհման ռեակցիաներին զուգահեռ տեղի են ունենում դրանց սինթեզի ռեակցիաներ։ Որոշ բակտերիաներ սպիտակուցներ են կազմում արտաքին սնուցման արդյունքում ստացված պատրաստի ամինաթթուներից, իսկ մյուս բակտերիաները ինքնուրույն սինթեզում են ամինաթթուները ազոտ և ածխածին պարունակող պարզ միացություններից։ Ամինաթթուների սինթեզը կարող է իրականացվել ամինացման, տրանսամինացիայի, ամիդացման, կարբոքսիլացման ռեակցիաներում։ Պրոկարիոտների մեծ մասը կարողանում է սինթեզել բոլոր ամինաթթուները, որոնք կազմում են բջջային սպիտակուցները։ Ամինաթթուների կենսասինթեզի առանձնահատկությունը սովորական կենսասինթետիկ ուղիների օգտագործումն է. տրիկարբոքսիլաթթվի ցիկլը, գլիկոլիզը, օքսիդատիվ պենտոզա-ֆոսֆատ ուղին և այլն: Ամինաթթուների սինթեզի հիմնական սկզբնական միացությունն է պիրուվատը և ֆումարատը:
ածխաջրերի նյութափոխանակությունայն տարբերվում է ավտոտրոֆներով և հետերոտրոֆներով (սխեմա 1): Ավտոտրոֆ բակտերիաները սինթեզում են բոլոր անհրաժեշտ ածխաջրերը ածխաթթու գազից։ Հետերոտրոֆ բակտերիաներում ածխաջրերի առաջացման հումքը կարող է լինել՝ (1) մեկ, երկու և երեք ածխածնային միացություններ. և (2) պոլիսախարիդներ (օսլա, գլիկոգեն, բջջանյութ): Վերջինիս պառակտման համար շատ հետերոտրոֆ բակտերիաներ ունեն էկզոֆերմենտներ (ամիլազ, պեկտինազ և այլն), որոնք հիդրոլիզացնում են պոլիսախարիդները՝ առաջացնելով գլյուկոզա, մալտոզ, ֆրուկտոզա և այլն։
Ավտոտրոֆ բակտերիաներում, Կալվինի ցիկլում, ածխածնի երկօքսիդից առաջանում է ռիբուլոզաֆոսֆատ-ֆոսֆոր-գլիցերինաթթու, որը ներառված է հակառակ ուղղությամբ ընթացող գլիկոլիզի ռեակցիաներում։ Հակադարձ սինթեզի վերջնական արդյունքը գլյուկոզան է:
Հետերոտրոֆ բակտերիաները գլյուկոզա են կազմում մեկ, երկու և երեք ածխածնային միացություններից՝ ներառելով դրանք նաև հակադարձ գլիկոլիզի ռեակցիայի մեջ։ Շնորհիվ այն բանի, որ գլիկոլիզի որոշ ռեակցիաներ անշրջելի են, հետերոտրոֆները ձևավորել են հատուկ ֆերմենտային ռեակցիաներ, որոնք թույլ են տալիս շրջանցել կատաբոլիկ ուղու անդառնալի ռեակցիաները։
Երբ պոլիսախարիդները ճեղքվում են հետերոտրոֆ բակտերիաների կողմից, ստացված դիսաքարիդները մտնում են բջիջներ և մալտոզայի, սախարոզայի և լակտոզայի ազդեցության տակ ենթարկվում են հիդրոլիզի և տրոհվում մոնոսաքարիդների, որոնք այնուհետև խմորվում կամ ներառվում են շաքարի փոխակերպման ռեակցիաներում:
լիպիդային նյութափոխանակություն. Ե՛վ էկզոգեն լիպիդները, և՛ ինտերստիցիալ նյութափոխանակության ամֆիբոլիտները կարող են ծառայել որպես սկզբնական նյութեր բակտերիաներում լիպիդների ձևավորման համար: Էկզոգեն լիպիդները ենթարկվում են բակտերիալ լիպազների և լիպոլիտիկ այլ ֆերմենտների: Բակտերիաների շատ տեսակներ կլանում են գլիցերինը, որը ծառայում է որպես պլաստիկ նյութի և էներգիայի աղբյուր։ Լիպիդների սինթեզի էնդոգեն աղբյուրները կարող են լինել ացետիլկոէնզիմ A, պրոպիոնիլ-APB, մալոնիլ-APB (ACP - ացետիլ-փոխադրող սպիտակուց), ֆոսֆոդիօքսիացետոն և այլն:
Զույգ թվով ածխածնի ատոմներով ճարպաթթուների սինթեզի սկզբնական սուբստրատը ացետիլկոենզիմ A է, կենտ թվով ածխածնի ատոմներով ճարպաթթուների համար՝ պրոպիոնիլ-APB և մալոնիլ-APB: Աերոբ պրոկարիոտներում թթվի մոլեկուլում կրկնակի կապերի առաջացումը տեղի է ունենում մոլեկուլային թթվածնի և դեզատուրազ ֆերմենտի մասնակցությամբ։ Անաէրոբ պրոկարիոտներում կրկնակի կապեր են ներմուծվում սինթեզի սկզբում՝ ջրազրկման ռեակցիայի արդյունքում։ Ֆոսֆոլիպիդների սինթեզի սկզբնական սուբստրատը ֆոսֆոդիօքսիացետոնն է (գլիկոլիտիկ ճանապարհի միջանկյալ միացություն), որի կրճատումը հանգեցնում է 3-ֆոսֆորոգլիցերինի առաջացմանը։ Այնուհետեւ վերջիններիս ավելացվում է 2 ճարպաթթվի մնացորդ՝ APB-ով կոմպլեքսի տեսքով։ Ռեակցիայի արտադրանքը ֆոսֆատիդային թթուն է, որի ակտիվացումը CTP-ով և հետագայում սերինի, ինոզիտոլի, գլիցերինի կամ այլ միացության ֆոսֆատային խմբին կցումը հանգեցնում է համապատասխան ֆոսֆոլիպիդների սինթեզին։
Միկրոօրգանիզմները, որոնք օքսոտրոֆ և հիպոտրոֆ են ճարպաթթուների համար (օրինակ՝ միկոպլազմաները) դրանք ստանում են պատրաստի վիճակում հյուրընկալող բջիջներից կամ սնուցող միջավայրից:
Մոնոնուկլեոտիդային փոխանակում. Պուրինի և պիրիմիդինի մոնոնուկլեոտիդները ԴՆԹ-ի և ՌՆԹ-ի կարևոր բաղադրիչներն են: Շատ պրոկարիոտներ ի վիճակի են ինչպես օգտագործել պատրաստի պուրինային և պիրիմիդինային հիմքերը, դրանց նուկլեոզիդներն ու նուկլեոտիդները, որոնք պարունակվում են սննդարար միջավայրում, այնպես էլ սինթեզել դրանք ցածր մոլեկուլային քաշի նյութերից: Բակտերիաներն ունեն ֆերմենտներ, որոնք կատալիզացնում են էկզոգեն պուրինի և պիրիմիդինի ածանցյալների փոխակերպումների հետևյալ փուլերը՝ ազոտային հիմք - նուկլեոզիդ - նուկլեոտիդ (մոնո - դի - - տրիֆոսֆատ):
Պուրինի և պիրիմիդինի մոնոնուկլեոտիդների սինթեզ de novoիրականացվում է անկախ եղանակներով։ Պուրինային նուկլեոտիդների սինթեզի ժամանակ հաջորդական ֆերմենտային ռեակցիաների արդյունքում առաջանում է ինոզինաթթու, որից պուրինային օղակի քիմիական փոփոխություններով սինթեզվում են ադենիլ (AMP) և գուանիլային (GMP) թթուները։ Պիրիմիդին նուկլեոտիդների սինթեզը սկսվում է օրոտիդիլաթթվի առաջացմամբ, որի դեկարբոքսիլացումից ստացվում է ուրիդիլաթթու (UMP): Վերջինից առաջանում է UTP, որի ացիլացումը հանգեցնում է CTP-ի առաջացմանը։
Դեզօքսիռիբոնուկլեոտիդներն առաջանում են դիֆոսֆատների կամ եռաֆոսֆատների մակարդակում համապատասխան ռիբոնուկլեոտիդների կրճատման արդյունքում։ ԴՆԹ-ին հատուկ նուկլեոտիդի՝ թիմիդիլաթթվի սինթեզը տեղի է ունենում դեզօքսյուրիդիլաթթվի ֆերմենտային մեթիլացման միջոցով։
Ion փոխանակում. Հանքային միացություններ՝ իոններ, NH 3 + , K + , Mg 2+ , Fe 2+ , SO 4 2- , PO 4 3- և այլ բակտերիաներ շրջակա միջավայրից ստացվում են ինչպես ազատ վիճակում, այնպես էլ այլ օրգանների հետ կապված վիճակում։ նյութեր. Կատիոնները և անիոնները բակտերիաների բջիջ են տեղափոխվում տարբեր ձևերով, որոնք նկարագրված են § 3-ում: Բակտերիաների բջիջ իոնների ներթափանցման արագության վրա ազդում են միջավայրի pH-ը և հենց միկրոօրգանիզմների ֆիզիոլոգիական ակտիվությունը:
Բ) բակտերիաների շնչառություն (էներգետիկ նյութափոխանակություն).
Կյանքի բոլոր գործընթացները կախված են էներգիայից, հետևաբար էներգիա ստանալը պրոկարիոտների նյութափոխանակության չափազանց կարևոր ասպեկտ է: Նրանք էներգիա են ստանում անաէրոբ և աերոբ շնչառությունից։
Շունչ, կամ կենսաբանական օքսիդացումն էլեկտրոնի փոխանցման կատաբոլիկ գործընթաց է դոնոր նյութից դեպի ընդունող նյութ, որն ուղեկցվում է մակրոէերգիկ միացություններում էներգիայի կուտակմամբ։ . Շնչառությունն իրականացվում է կատաբոլիկ ռեակցիաների գործընթացում, որի արդյունքում բարդ օրգանական նյութերը, տրոհվելով, էներգիա են տալիս և վերածվում պարզ միացությունների։ Մակրոէերգիկ նյութերում (ATP, GTP, UTP և այլն) կուտակված էներգիան օգտագործվում է անաբոլիկ ռեակցիաներում։
Ըստ շնչառության ձևի՝ միկրոօրգանիզմները դասակարգվում են պարտավորեցնող (խիստ) աերոբներ, պարտավորեցնող անաէրոբներև ֆակուլտատիվ անաէրոբներ.
պարտադիր աերոբներանհրաժեշտ է անվճար թթվածին. Օրգանական միացությունները (ածխաջրեր, ճարպեր, սպիտակուցներ) էլեկտրոնների դոնորներ են մարդու ախտածին աերոբ-քիմոօրգանոտրոֆներում, իսկ մոլեկուլային թթվածինը էլեկտրոն ընդունող է: Էներգիայի կուտակումը ATP-ի տեսքով քիմոօրգանոտրոֆ աերոբներում տեղի է ունենում էլեկտրոն դոնորների օքսիդատիվ ֆոսֆորիլացման ժամանակ։ Աերոբներն ունեն ցիտոքրոմներ (մասնակցում են էլեկտրոնների փոխանցմանը), ինչպես նաև ֆերմենտներ (կատալազ, սուպերօքսիդ դիսմուտազ, պերօքսիդազ), որոնք անակտիվացնում են շնչառության ընթացքում առաջացած թունավոր թթվածնային ռադիկալները։ Սուպերօքսիդ դիսմուտազն ապաակտիվացնում է ամենաթունավոր մետաբոլիտը՝ H 2 O 2-ում առկա սուպերօքսիդ ռադիկալը: Կատալազ ֆերմենտը H 2 O 2-ը վերածում է H 2 O և O 2-ի:
Աերոբների հատուկ խումբ են միկրոաերոֆիլ բակտերիաներ, որոնք, թեև էներգիայի համար թթվածնի կարիք ունեն, բայց ավելի լավ են զարգանում CO 2-ի ավելացված պարունակությամբ, օրինակ՝ սեռի բակտերիաները Campylobacterև Helicobacter.
պարտադիր անաէրոբներազատ թթվածնի կարիք չունեն, ընդհակառակը, նույնիսկ փոքր քանակությամբ թթվածինը թունավոր ազդեցություն ունի նրանց վրա։ Մարդկանց ախտածին անաէրոբ-քիմոօրգանոտրոֆների էլեկտրոնների դոնորները տարբեր օրգանական միացություններ են (հիմնականում ածխաջրեր): Քիմիօրգանոտրոֆ անաէրոբներում էլեկտրոնների ընդունիչը օրգանական թթվածին պարունակող միացություններ են՝ թթուներ կամ կետոններ, այսինքն՝ էլեկտրոն ընդունողը թթվածին է, կապված օրգանական հատվածի հետ։ Այս պրոկարիոտներում էներգիայի կուտակումը տեղի է ունենում սուբստրատի ֆոսֆորիլացման ժամանակ։ Պարտադիր անաէրոբները, որպես կանոն, չունեն ցիտոքրոմներ և ֆերմենտներ, որոնք ապաակտիվացնում են թթվածնային ռադիկալները (կատալազ- և սուպերօքսիդ դիսմուտազային բացասական):
Քիմոլիտոտրոֆ անաէրոբներում, որոնք մարդու համար պաթոգեն չեն, էլեկտրոն ընդունողն անօրգանական թթվածին պարունակող միացություններն են՝ նիտրատներ, սուլֆատներ, կարբոնատներ:
Անաէրոբների հատուկ խումբ են հանդիսանում աերոտոլերանտ բակտերիաները, որոնք ունակ են աճել մթնոլորտային թթվածնի առկայության դեպքում, բայց չեն օգտագործում այն որպես էլեկտրոն ընդունող (օրինակ՝ կաթնաթթվային բակտերիաներ)։ Աերոտոլերանտ կատալազ և սուպերօքսիդ դիսմուտազ պրոկարիոտները դրական են:
Ֆակուլտատիվ անաէրոբներկարող է գոյություն ունենալ ինչպես թթվածնային, այնպես էլ անօքսիկ միջավայրում: Նրանց էլեկտրոնների դոնորները օրգանական նյութեր են. Էլեկտրոնների ընդունիչները, կախված շրջակա միջավայրի պայմաններից, մոլեկուլային կամ թթվածին են կապված օրգանական և անօրգանական միացություններում: Ֆակուլտատիվ անաէրոբները կարող են էներգիա կուտակել ինչպես օքսիդատիվ, այնպես էլ սուբստրատի ֆոսֆորիլացման ժամանակ: Ինչպես աերոբները, բակտերիաների այս խումբն ունի ցիտոքրոմներ և հակաօքսիդանտ պաշտպանական ֆերմենտներ:
Էներգիայի ստացման հիմնական սուբստրատը ածխաջրերն են, որոնք շնչառության տարբեր տեսակների քիմեմետերոտրոֆ պրոկարիոտներում կարող են կատաբոլիզացվել ացետիլկոենզիմ A-ի («ակտիվացված քացախաթթու»): Լիպիդները և սպիտակուցները կարող են հանդես գալ որպես էներգիայի սուբստրատներ, քանի որ ացետիլ կոենզիմ A-ն նույնպես նրանց նյութափոխանակության միջանկյալ արտադրանքներից է (սխեմա 2):
Ածխաջրերի կատաբոլիզմը քիմոօրգանոտրոֆ պրոկարիոտներում ներառում է. բ) աերոբ պրոցես - եռաքարբոքսիլաթթվի ցիկլ (Կրեբսի ցիկլ): Անաէրոբ պրոցեսները տեղի են ունենում բոլոր պրոկարիոտների մոտ, մինչդեռ աերոբ գործընթացը բնորոշ է միայն պարտադիր աերոբներին և ֆակուլտատիվ անաէրոբներին: Անաէրոբ ուղիները հիմնված են սուբստրատի ֆոսֆորիլացման վրա, մինչդեռ աերոբ գործընթացները հիմնված են օքսիդատիվ ֆոսֆորիլացման վրա:
Հասկացությունների սահմանում.
Ստերիլիզացումը, ախտահանումը և հակասեպտիկ միջոցները ժամանակակից բժշկական և հատկապես վիրաբուժական պրակտիկայի անբաժանելի մասն են: Այս մեթոդների սկզբունքները և գործնական կիրառումը կարևոր է, քանի որ շատ պոտենցիալ պաթոգեն միկրոօրգանիզմներ կարող են երկար ժամանակ գոյատևել հյուրընկալող տարածքից դուրս, ցուցաբերել բարձր դիմադրություն ֆիզիկական և քիմիական ախտահանիչների նկատմամբ և համեմատաբար հեշտությամբ տարածվել մարդուց մարդ:
Հակասեպտիկներ- քիմիական մեթոդներով պաթոգեն կամ օպորտունիստական միկրոօրգանիզմների աճի ոչնչացում կամ կանխարգելում. Այս տերմինը սովորաբար օգտագործվում է կենդանի հյուսվածքների վրա քիմիական պատրաստուկի արտաքին կիրառման համար:
հակասեպտիկ- նյութ, որը արգելակում է աճը կամ ոչնչացնում միկրոօրգանիզմը (առանց բակտերիաների սպորների վրա գործողության): Տերմինը հատուկ է այն նյութերին, որոնք օգտագործվում են կենդանի հյուսվածքների վրա տեղական ազդեցության համար:
Ասեպսիսնշանակում է ս sepsis-ի բացակայություն, բայց ընդհանուր առմամբ այս տերմինն օգտագործվում է որևէ կենդանի օրգանիզմի բացակայությունն ընդգծելու համար։ Ասեպտիկ մեթոդներ նշանակում է ցանկացած ընթացակարգ, որը նախատեսված է կենդանի օրգանիզմների ոչնչացման և դրանցով նորից աղտոտումը կանխելու համար: Ժամանակակից վիրաբուժական և մանրէաբանական մեթոդները հիմնված են ասեպտիկ պրոցեդուրաների վրա:
Կենսասպանություն- նյութ, որը սպանում է բոլոր կենդանի միկրոօրգանիզմներին, ինչպես պաթոգեն, այնպես էլ ոչ ախտածին, ներառյալ սպորները:
Բիոստատ- գործակալ, որը կանխում է միկրոօրգանիզմների աճը, բայց պարտադիր չէ, որ սպանի նրանց:
Ախտահանում- միկրոօրգանիզմների հեռացում առանց քանակական որոշման. Այս տերմինը հարաբերական է. Մանրէների վերջնական հեռացումը կարող է իրականացվել մանրէազերծման կամ ախտահանման միջոցով:
Ախտահանում- գործընթաց, որը նվազեցնում կամ վերացնում է բոլոր պաթոգենները, բացի սպորներից:
մանրէասպան- նյութ, որը ոչնչացնում է միկրոօրգանիզմները, հատկապես պաթոգենները: Գերմիցիդը չի ոչնչացնում սպորները:
Սանացիա- մեթոդ, որով մանրէաբանական աղտոտվածությունը նվազեցվում է «անվտանգ» մակարդակի: Այս մեթոդը նախկինում կիրառվում էր անշունչ առարկաները «մաքրելու» համար։
Ստերիլիզացում- ֆիզիկական գործոնների և (կամ) քիմիական նյութերի օգտագործումը մանրէաբանական կյանքի բոլոր ձևերի ամբողջական ոչնչացման կամ ոչնչացման համար.
Ստերիլիզացում.
Ստերիլիզացումը սահմանվում է որպես բոլոր միկրոօրգանիզմների և դրանց սպորների ոչնչացում կամ հեռացում (ֆիլտրման միջոցով): Ստերիլիզացումը սովորաբար իրականացվում է ջերմության միջոցով: Մանրէազերծումը, լինելով մանրէաբանական լաբորատորիայի աշխատանքի առօրյաներից մեկը, կարևոր մեթոդ է ապահովելու, որ մշակույթները, սարքավորումները, պարագաներն ու կրիչները նպաստեն միայն անհրաժեշտ միկրոօրգանիզմների աճին, իսկ մյուս մանրէները ոչնչացվեն: Կան մանրէազերծման այդպիսի տեսակներ՝ այրիչի կրակի մեջ կալցինացիա, եռում, հոսող գոլորշիով գործողություն, գոլորշու ճնշման տակ ավտոկլավում, չոր ջերմություն, պաստերիզացում, թինդալիզացիա, քիմիական, սառը (մեխանիկական) ստերիլիզացում։
Ստերիլիզացման մեթոդների ընտրություն.
Ստերիլիզացման մեթոդներ ընտրելիս պետք է հաշվի առնել հետևյալ պահանջները.
1. Գործունեություն՝ մանրէասպան, սպորիցիդ, տուբերկուլյոզ, ֆունգիցիդ և վիրուսասպան։
2. Ընթացակարգի արագությունը. ստերիլիզացումը պետք է իրականացվի հնարավորինս արագ:
3. Անթափանցելիություն. մանրէազերծող նյութերը պետք է կարողանան ներթափանցել փաթեթավորման միջով և գործիքի ներս:
4. Համատեղելիություն. մի քանի անգամ մանրէազերծված նյութերի կառուցվածքի կամ ֆունկցիայի փոփոխություն չպետք է լինի:
5. Ոչ թունավոր. չպետք է սպառնալիք լինի մարդու առողջությանը և շրջակա միջավայրին:
6. Օրգանական նյութի կայունությունը. ստերիլիզացման արդյունավետությունը չպետք է նվազեցվի օրգանական նյութի առկայության դեպքում:
7. Հարմարվողականություն. ստերիլիզացված նյութի մեծ և փոքր ծավալների համար օգտագործելու ունակություն:
8. Ժամանակի վերահսկում. մշակման ցիկլը պետք է հեշտությամբ և ճշգրիտ վերահսկվի:
9. Գինը` սարքավորումների, տեղադրման և շահագործման ողջամիտ արժեքը:
 Ֆիզիկական ստերիլիզատորներ
Ֆիզիկական ստերիլիզատորներ
թաց ջերմորեն,որը ձևավորվում է գոլորշու ավտոկլավացման գործընթացում, հիմնական մանրէազերծող նյութն է, որն օգտագործվում է կլինիկական մանրէաբանական լաբորատորիաներում: Ավտոկլավներն օգտագործվում են մշակութային միջավայրերի, հրակայուն նյութերի ստերիլիզացման և վարակիչ թափոնների բուժման համար: Գոլորշի մանրէազերծիչը կամ ավտոկլավը մեկուսացված ճնշման խցիկ է, որն օգտագործում է հագեցած գոլորշի բարձր ջերմաստիճան առաջացնելու համար (Նկար 1): Օդը խցիկից հեռացվում է զանգվածային տեղաշարժով կամ վակուումով: Առավել հաճախ օգտագործվող ավտոկլավները՝ ըստ քաշի փոխարինման: Ավելի թեթև գոլորշի է մտցվում խցիկ՝ ավելի ծանր օդը տեղափոխելու համար: Սեղմված գոլորշու կարճատև ազդեցությունը կարող է ոչնչացնել բակտերիաների սպորները: Կուլտուրայի միջավայրի և այլ նյութերի սովորական մանրէազերծման համար ազդեցության ժամանակը 15 րոպե է 121°C ջերմաստիճանում և 1 քառակուսի սանտիմետրում 1,5 կգ ճնշում: Վարակիչ թափոնների դեպքում ազդեցության ժամանակը ավելացվում է մինչև 30-60 րոպե: Բացի ճիշտ ժամանակից և ջերմաստիճանից, ստերիլիզացման համար շատ կարևոր է գոլորշու հետ անմիջական շփումը: Վարակիչ նյութերի հետ աշխատելիս պետք է ապահովվի գոլորշու առավելագույն ներթափանցումը թափոնների մեջ: Նման նյութը պետք է մշակվի 132ºС ջերմաստիճանում: Հականեոպլաստիկները, թունավոր քիմիկատները և ռադիոիզոտոպները, որոնք չեն կարող քայքայվել, և անկայուն քիմիական նյութերը չպետք է ավտոկլավացվեն, քանի որ դրանք կարող են գոլորշիանալ և տարածվել խցիկում, երբ ենթարկվում են ջերմության:
 Չոր ջերմային ստերիլիզացումօգտագործվում է այն նյութերի համար, որոնք հնարավոր չէ ստերիլիզացնել գոլորշու միջոցով՝ վնասվելու հնարավորության կամ նյութի գոլորշու նկատմամբ անթափանցելիության պատճառով։ Չոր ջերմությունը ավելի քիչ արդյունավետ է, քան խոնավ ջերմությունը և պահանջում է ավելի երկար ազդեցության ժամանակ և ավելի բարձր ջերմաստիճան: Չոր ջերմությամբ ստերիլիզացումը սովորաբար իրականացվում է չոր ջերմային պահարանում (նկ. 2): Չոր ջերմային ստերիլիզացման մեխանիզմը օքսիդատիվ գործընթաց է։ Նյութերի օրինակներ, որոնց համար օգտագործվում է չոր ջերմային ստերիլիզացում, յուղեր, փոշիներ, սուր գործիքներ և ապակյա իրեր են: Որպես վարակիչ թափոնների բուժման այլընտրանքային մեթոդներ օգտագործվում են չոր ջերմությունը կամ ջերմային ապաակտիվացում-ստերիլիզացումը։
Չոր ջերմային ստերիլիզացումօգտագործվում է այն նյութերի համար, որոնք հնարավոր չէ ստերիլիզացնել գոլորշու միջոցով՝ վնասվելու հնարավորության կամ նյութի գոլորշու նկատմամբ անթափանցելիության պատճառով։ Չոր ջերմությունը ավելի քիչ արդյունավետ է, քան խոնավ ջերմությունը և պահանջում է ավելի երկար ազդեցության ժամանակ և ավելի բարձր ջերմաստիճան: Չոր ջերմությամբ ստերիլիզացումը սովորաբար իրականացվում է չոր ջերմային պահարանում (նկ. 2): Չոր ջերմային ստերիլիզացման մեխանիզմը օքսիդատիվ գործընթաց է։ Նյութերի օրինակներ, որոնց համար օգտագործվում է չոր ջերմային ստերիլիզացում, յուղեր, փոշիներ, սուր գործիքներ և ապակյա իրեր են: Որպես վարակիչ թափոնների բուժման այլընտրանքային մեթոդներ օգտագործվում են չոր ջերմությունը կամ ջերմային ապաակտիվացում-ստերիլիզացումը։
Պաստերիզացիան ոչնչացնում է պաթոգենները՝ նյութը 15 վայրկյան արագ տաքացնելով մինչև 71,1°C, որին հաջորդում է արագ սառեցումը: Պաստերիզացումը ստերիլիզացում չէ, քանի որ ոչ բոլոր միկրոօրգանիզմներն են դրա նկատմամբ զգայուն: Այս մեթոդը վերացրեց սննդային ճանապարհով այնպիսի հիվանդությունների փոխանցումը, ինչպիսիք են ստամոքս-աղիքային տուբերկուլյոզը և Քյու տենդը:
Tyndalizationջերմային ստերիլիզացման ընդհատվող մեթոդ է, որը կարող է օգտագործվել լուծույթներում առկա բոլոր բակտերիաների ոչնչացման համար: Քանի որ աճող բակտերիաները հեշտությամբ ոչնչացվում են կարճ եռալով (5 անգամ 1 ժամվա ընթացքում 5 րոպեի ընթացքում), այն ամենը, ինչ պետք է արվի, այն է, որ լուծույթը որոշ ժամանակ կանգնի, մինչև ջերմությունը կխախտի սպորների հասունացումը: ջերմության նկատմամբ նրանց դիմադրության զգալի կորուստ:
Զտումգործընթաց է, որն օգտագործվում է լուծույթներից, օդից և այլ գազերից մանրէները և մանրադիտակային մասնիկները հեռացնելու համար: Լաբորատորիայում ֆիլտրացիայի միջոցով ստերիլիզացման ամենատարածված օգտագործումը ախտորոշման, կուլտուրայի միջավայրի, հյուսվածքների կուլտուրայի միջավայրի, շիճուկների, շիճուկի բաղադրիչներ պարունակող լուծույթների մշակումն է: Զտման մեկ այլ տարածված կիրառություն օդի և գազերի մանրէազերծումն է: Պլաստիկ կամ թղթե թաղանթային ֆիլտրերը, որոնք տարբերվում են ծակոտիների տրամագծով (մոտ 12-ից մինչև 0,22 մկմ) և օգտագործվում են մեխանիկական բաժանման համար, նաև ծառայում են միկրոսկոպիկ հետազոտության համար հեղուկներից մանրէներ հավաքելու կամ անմիջապես ֆիլտրի վրա մշակելու համար, երբ տեղադրվում են մակերեսային թրջված սննդանյութի վրա։ միջին.
ուլտրամանուշակագույն ճառագայթումէլեկտրամագնիսական ալիքային ճառագայթման տեսակ է, որը գործում է բջջային նուկլեինաթթվի վրա։ Միկրոօրգանիզմները խիստ զգայուն են 254 նմ ալիքի երկարությամբ ուլտրամանուշակագույն ճառագայթների ազդեցության նկատմամբ։ Ուլտրամանուշակագույն լույսը առավել լայնորեն օգտագործվում է օդում կամ մակերեսների վրա միկրոօրգանիզմները ոչնչացնելու համար: Այլ կիրառումներ են որոշ քիմիական նյութերի և պլաստմասսաների սառը մանրէազերծումը դեղագործական կիրառությունների համար, շիճուկի մանրէազերծումը բջջային կուլտուրաների համար և ջրի ախտահանումը: Ուլտրամանուշակագույն ճառագայթման որպես մանրէազերծիչի զգալի թերությունը նյութերի մեջ ներթափանցելու անկարողությունն է:
իոնացնող ճառագայթումէլեկտրամագնիսական սպեկտրում այն մահացու ազդեցություն է ունենում միկրոօրգանիզմների վրա։ Այս սպեկտրը ներառում է միկրոալիքներ, γ-ճառագայթներ, ռենտգենյան ճառագայթներ և էլեկտրոնների հոսք: Իոնացնող ճառագայթման մահացու ազդեցությունը տեղի է ունենում թիրախային մոլեկուլի վրա անմիջական գործողության պատճառով, որի արդյունքում էներգիան փոխանցվում է մոլեկուլին. իսկ անուղղակի գործողության շնորհիվ՝ ռադիկալների դիֆուզիոն։
ուլտրաձայնային էներգիացածր հաճախականությամբ անակտիվացնում է միկրոօրգանիզմները ջրային լուծույթներում: Ֆիզիկական ազդեցությունը sonication պայմանավորված է cavitation. Ուլտրաձայնային մաքրող միջոցները և այլ սարքերը հաճախ օգտագործվում են գործիքները մաքրելու համար, բայց չեն համարվում ստերիլիզատորներ: Այնուամենայնիվ, ուլտրաձայնի համադրությունը քիմիական բուժման հետ սպանում է միկրոօրգանիզմները:
Քիմիական ստերիլիզատորներ
2 % գլյուտարալդեհիդՈրպես հեղուկ քիմիական ստերիլիզատոր, այն նախկինում լայնորեն օգտագործվում էր բժշկական և վիրաբուժական նյութերի մշակման համար, որոնք հնարավոր չէ մանրէազերծել տաքացման կամ ճառագայթման միջոցով: Գլյուտարալդեհիդը նույնպես օգտագործվում է պատվաստանյութերի պատրաստման մեջ։
Ախտահանում.
Ախտահանումը կարող է իրականացվել քիմիական մեթոդներով կամ եռալով։ Եռացումը արդյունավետ մեթոդ է այնպիսի գործիքների ախտահանման համար, ինչպիսիք են ասեղները և ներարկիչները, եթե ավտոկլավը հասանելի չէ: Նախապես մաքրված բժշկական գործիքները պետք է եփել 20 րոպե։ Քիմիական ախտահանումն օգտագործվում է ջերմային զգայուն սարքավորումների համար, որոնք կարող են վնասվել բարձր ջերմաստիճանից: Քիմիական ախտահանիչ միջոցները, ինչպիսիք են քլորի բաղադրիչները, էթիլային և իզոպրոպիլային ալկոհոլը, չորրորդական ամոնիումի բաղադրիչները և գլյուտարալդեհիդը լայնորեն օգտագործվում են:
Քիմիական ախտահանիչներ.
Ալկոհոլ (էթիլ և իզոպրոպիլ), ջրում լուծված 60-85% կոնցենտրացիայով շատ արդյունավետ է ախտահանման համար։ Սպիրտները մանրէասպան, ֆունգիցիդ և տուբերկուլյոզ են, բայց չեն ազդում սպորների վրա: Էթիլային սպիրտն ունի վիրուսային ակտիվության ավելի լայն սպեկտր, քան իզոպրոպիլային ալկոհոլը, ուստի այն ավելի արդյունավետ է լիպոֆիլ և հիդրոֆիլ վիրուսների դեմ:
Լուծում 37% ֆորմալդեհիդֆորմալին, որը կոչվում է ֆորմալին, կարող է օգտագործվել որպես ստերիլիզատոր, մինչդեռ դրա 3-8% կոնցենտրացիաները կարող են օգտագործվել որպես ախտահանիչ:
Ֆենոլիր մաքուր տեսքով չի օգտագործվում որպես ախտահանիչ՝ իր թունավորության, ուռուցքների զարգացում առաջացնելու ունակության և կոռոզիայի պատճառով: Ֆենոլի ածանցյալները, որոնցում ֆունկցիոնալ խումբը (քլոր, բրոմ, ալկիլ, բենզիլ, ֆենիլ, ամիլ) փոխարինում է անուշաբույր օղակի ջրածնի ատոմներից մեկին, լայնորեն օգտագործվում են որպես ախտահանիչներ։ Այս փոխարինումը նվազեցնում է ֆենոլի թերությունները: Ֆենոլի բաղադրիչները սպանում են մանրէները ֆերմենտային համակարգերի ապաակտիվացման, սպիտակուցների տեղումների և բջջային պատի և թաղանթի խզման պատճառով: Սովորաբար օգտագործվում են 2-5% կոնցենտրացիաներ, ավելի ցածր կոնցենտրացիաները պահանջում են ավելի երկար ազդեցություն:
Հալոգեններ.Լաբորատոր պրակտիկայում ախտահանման համար օգտագործվում են միայն քլոր և յոդ: Շնորհիվ այն բանի, որ քլորը հզոր օքսիդացնող նյութ է, ենթադրվում է, որ այն սպանում է մանրէները օքսիդացման միջոցով: Ենթադրվում է, որ յոդը սպանում է միկրոօրգանիզմներին՝ արձագանքելով ամինաթթուների N-H և S-H խմբերի, ինչպես նաև ամինաթթվի թիրոզինի և չհագեցած ճարպաթթուների ածխածին-ածխածին կրկնակի կապերի ֆենոլային խմբի հետ։ Սովորական բուժումը ներառում է 2-5% ֆորմալդեհիդի լուծույթի ցողում գոլորշու առկայության դեպքում 60-80ºC ջերմաստիճանում:
Հակասեպտիկներ.
Հակասեպտիկները կարելի է գտնել մանրէաբանական լաբորատորիաներում, հիմնականում այն նյութերում, որոնք օգտագործվում են ձեռքերը լվանալու համար: Այն դեպքերում, երբ բժշկական անձնակազմը շտապ օգնություն է ցուցաբերում հիվանդներին, ովքեր օգտագործում են հակաբակտերիալ նյութեր պարունակող նյութեր, դա նվազեցնում է հիվանդանոցային վարակների թիվը: Ձեռքերի լվացման մեջ հայտնաբերված ամենատարածված քիմիական նյութերն են սպիրտները, քլորիխիդին գլյուկոնատը, յոդոֆորները, քլորօքսիլենոլը և տրիկլոզանը:
Թափոնների և աղբի մշակման ավանդական մեթոդներն են այրումը և գոլորշու ստերիլիզացումը:
Այրվողթափոնների և բեկորների հետ աշխատելու ընտրության մեթոդն է: Այս մեթոդը աղբը դարձնում է ոչ վարակիչ, ինչպես նաև փոխում է դրա ձևն ու չափը։ Ստերիլիզացումը թափոնների մաքրման արդյունավետ մեթոդ է, սակայն այն չի փոխում իր ձևը։ Գոլորշիով մանրէազերծումը ավտոկլավում 121°C ջերմաստիճանում առնվազն 15 րոպե տևողությամբ ոչնչացնում է մանրէների կյանքի բոլոր ձևերը, ներառյալ մեծ թվով բակտերիաների սպորները: Այս տեսակի ամբողջական ստերիլիզացումը կարող է իրականացվել նաև չոր ջերմության միջոցով 160-170ºC ջերմաստիճանում 2-4 ժամ: Այնուամենայնիվ, պետք է ապահովել, որ չոր ջերմությունը շփվի ստերիլիզացման ենթակա նյութի հետ: Հետևաբար, հեղուկ պարունակող շշերը պետք է փակված լինեն խցաններով կամ բամբակյա շվաբրերով, որպեսզի գոլորշին և ջերմությունը փոխանակվեն շշերի օդի հետ: Թափոններ պարունակող կենսավտանգների տարաները պետք է սերտորեն կապված լինեն: Մանրէազերծված կենսավտանգավոր նյութերը պետք է փակվեն համապատասխան պիտակավորված տարաներում:
Գոլորշով ստերիլիզացում (ավտոկլավ):Վարակիչ աղբը համարվում է վարակազերծված, երբ վեգետատիվ բակտերիաների, սնկերի, միկոբակտերիաների և լիպիդ պարունակող վիրուսների թիվը նվազում է 6 լգ անգամ, իսկ բակտերիալ էնդոսպորները՝ 4 լգ անգամ:
Բակտերիաների սնուցում.
Սնուցում. Բակտերիալ բջջի սնուցման ներքո պետք է հասկանալ փոխակերպման ռեակցիաների արդյունքում պլաստիկ նյութի և էներգիայի կլանման և յուրացման գործընթացը: . Պրոկարիոտների սնուցման տեսակները բարդ են և բազմազան։ Դրանք տարբերվում են՝ կախված բակտերիաների բջիջ սնուցիչների ներթափանցման եղանակից, ածխածնի և ազոտի աղբյուրներից, էներգիայի ստացման եղանակից և էլեկտրոնների դոնորների բնույթից։
սննդանյութերի տեղափոխում բջիջկարող է իրականացվել 3 մեխանիզմներով՝ պասիվ դիֆուզիա, հեշտացված դիֆուզիա և ակտիվ տրանսպորտ։
պասիվ դիֆուզիաէներգիայից կախված ոչ սպեցիֆիկ գործընթաց է, որն իրականացվում է նյութերի կոնցենտրացիայի գրադիենտի երկայնքով (ավելի բարձր կոնցենտրացիա ունեցող միջավայրից նյութը պասիվորեն, ըստ օսմոսի օրենքների, մտնում է ավելի ցածր կոնցենտրացիայով միջավայր): Պասիվ դիֆուզիայի միջոցով մանրէների բջիջ են մտնում սահմանափակ քանակությամբ նյութեր, որոշ իոններ, մոնոսաքարիդներ։ Պասիվ դիֆուզիայի ժամանակ նյութերի փոխանցման արագությունը աննշան է և կախված է փոխադրվող մոլեկուլների լիպոֆիլությունից և չափից։
Հեշտացված դիֆուզիոննյութերի չցնդող փոխադրում է կոնցենտրացիայի գրադիենտով պերմեազային ֆերմենտների օգնությամբ։ Պերմեազները հատուկ թաղանթային սպիտակուցներ են, որոնք հեշտացնում են նյութերի անցումը ցիտոպլազմային թաղանթով։ Պերմեազն իր վրա ամրացնում է փոխանցված նյութի մոլեկուլը, որի հետ միասին հաղթահարում է ցիտոպլազմային թաղանթը, որից հետո «նյութ-պերմեազ» կոմպլեքսը տարանջատվում է։ Ազատված պերմեազը օգտագործվում է այլ մոլեկուլներ անցկացնելու համար: Պրոկարիոտների մոտ միայն գլիցերինը բջիջ է մտնում հեշտացված դիֆուզիայի միջոցով: Այս դեպքում գլիցերինի ներբջջային կոնցենտրացիան համապատասխանում է բջիջից դուրս: Հեշտացված դիֆուզիան առավել բնորոշ է էուկարիոտիկ միկրոօրգանիզմներին:
ակտիվ տրանսպորտ- սա նյութերի էներգիայից կախված փոխանցում է բջիջ՝ ընդդեմ կոնցենտրացիայի գրադիենտի՝ հատուկ ֆերմենտների օգնությամբ: Նյութերի ճնշող մեծամասնությունը (իոններ, ածխաջրեր, ամինաթթուներ, լիպիդներ և այլն) ակտիվ տրանսպորտով մտնում են բակտերիաների բջիջ։ Ակտիվ փոխադրումը կարող է իրականացվել՝ (1) առանց տեղափոխվող նյութի քիմիական փոփոխության. (2) քիմիական փոփոխությամբ:
Բակտերիաները բջջային կառուցվածքի պրոկարիոտ միկրոօրգանիզմներ են։ Նրանց չափերը 0,1-ից մինչև 30 մկմ են: Մանրէները չափազանց տարածված են: Նրանք ապրում են հողում, օդում, ջրում, ձյան և նույնիսկ տաք աղբյուրներում, կենդանիների մարմնի վրա, ինչպես նաև կենդանի օրգանիզմների, այդ թվում՝ մարդու մարմնի ներսում։
Բակտերիաների բաշխումը տեսակների վրա հիմնված է մի քանի չափանիշների վրա, որոնց թվում առավել հաճախ հաշվի են առնվում միկրոօրգանիզմների ձևը և դրանց տարածական բաշխումը: Այսպիսով, ըստ բջիջների ձևի, բակտերիաները բաժանվում են.
Coci - միկրո-, դիպլո-, strepto-, staphylococci, ինչպես նաև սարկիններ;
Rod-shaped - monobacteria, diplobacteria եւ streptobacteria;
Խճճված տեսակներ - վիբրիոներ և սպիրոխետներ:
Bergey-ի որոշիչը համակարգում է բոլոր հայտնի բակտերիաները՝ ըստ բակտերիաների նույնականացման սկզբունքների, որոնք ամենալայն տարածում են գտել պրակտիկ մանրէաբանության մեջ՝ հիմնվելով բջջային պատի կառուցվածքի տարբերությունների վրա և կապված Գրամ ներկերի հետ: Բակտերիաների նկարագրությունը տրվում է ըստ խմբերի (բաժինների), որոնք ներառում են ընտանիքներ, սեռեր և տեսակներ. որոշ դեպքերում խմբերը ներառում են դասեր և պատվերներ: Մարդկանց համար պաթոգեն բակտերիաները ընդգրկված են փոքր թվով խմբերում:
Բանալին առանձնացնում է բակտերիաների չորս հիմնական կատեգորիաներ.
Gracillicutes [լատ. gracilis, նազելի, նիհար, + cutis, մաշկ] - տեսակ բարակ բջջային պատով, ներկված։ գրամ բացասական;
firmicutes [լատ. flrmus, ուժեղ, + cutis, skin] - բակտերիաներ հաստ բջջային պատով, ներկում գրամ դրական;
Tenericutes [լատ. նուրբ, նուրբ, + կնճիռ, մաշկ] - բակտերիաներ, որոնք չունեն բջջային պատ(միկոպլազմաներ և Mollicutes դասի այլ ներկայացուցիչներ)
Mendosicutes [լատ. mendosus, անկանոն, + cutis, մաշկ] - արխեբակտերիաներ (մեթան և սուլֆատ վերականգնող, հալոֆիլ, ջերմաֆիլ և արխեբակտերիաներ, որոնք զուրկ են բջջային պատից):
Խումբ 2 Burgey-ի որոշիչ. Աերոբիկ և միկրոաերոֆիլ շարժական ոլորված և կորացած գրամ-բացասական բակտերիաներ: Մարդկանց համար ախտածին տեսակները ներառված են Campylobacter, Helicobacters Spirillum ցեղերի մեջ։
Բերգեյի որոշիչի 3-րդ խումբ. Ոչ շարժուն (հազվադեպ շարժուն) Գրամ-բացասական բակտերիաներ: Չի պարունակում ախտածին տեսակներ։
Burgey-ի որոշիչի 4-րդ խումբ. Գրամ-բացասական աերոբիկ և միկրոաերոֆիլ ձողեր և կոկիներ: Մարդկանց համար ախտածին տեսակներն ընդգրկված են Legionellaceae, Neisseriaceae և Pseudomonada-ceae ընտանիքներում, խումբը ներառում է նաև Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Francisella, Kingella և Moraxela սեռերի պաթոգեն և պատեհապաշտ բակտերիաները:
Բերգեյի որոշիչի 5-րդ խումբ. Ֆակուլտատիվ անաէրոբ Գրամ-բացասական ձողեր: Խումբը կազմված է երեք ընտանիքից՝ Enterobacteriaceae, Vibrionaceae և Pasteurellaceae, որոնցից յուրաքանչյուրը ներառում է պաթոգեն տեսակներ, ինչպես նաև Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella և Streptobacillus սեռերի պաթոգեն և պատեհապաշտ բակտերիաներ։
Բերգեյի որոշիչի 6-րդ խումբ. Գրամ-բացասական անաէրոբ ուղիղ, կոր և պարուրաձև բակտերիաներ: Պաթոգեն և պատեհապաշտ տեսակներն ընդգրկված են Bacteroides, Fusobacterium, Porphoromonas և Prevotelta ցեղերում:
Բերգեյի որոշիչի 7-րդ խումբ. Բակտերիաները, որոնք կատարում են սուլֆատի կամ ծծմբի դիսիմիլացիոն նվազեցում: Չի ներառում ախտածին տեսակները:
Բերգեյի որոշիչի 8-րդ խումբ. Անաէրոբ գրամ-բացասական կոկի. Ներառում է Veillonella սեռի օպորտունիստական բակտերիաները։
Բերգեյի որոշիչի 9-րդ խումբ. Ռիկեցիա և քլամիդիա. Երեք ընտանիք՝ Rickettsiaceae, Bartonellaceae և Chlamydiaceae, որոնցից յուրաքանչյուրը պարունակում է մարդկանց համար ախտածին տեսակներ:
Burgey-ի ուղեցույցի 10-րդ և 11-րդ խմբերը ներառում են անօքսի- և թթվածին ֆոտոտրոֆ բակտերիաներ, որոնք պաթոգեն չեն մարդկանց համար:
Burgey-ի որոշիչի 12-րդ խումբ. Աերոբ քիմոլիտոտրոֆ բակտերիաներ և հարակից օրգանիզմներ: Այն միավորում է ծծմբի-երկաթի և մանգանի օքսիդացնող և նիտրացնող բակտերիաները, որոնք վնաս չեն հասցնում մարդկանց:
Burgey-ի ուղեցույցի 13-րդ և 14-րդ խմբերը ներառում են բողբոջող և/կամ աճող բակտերիաներ և թաղանթ ձևավորող բակտերիաներ: Ներկայացված է ազատ ապրող, մարդկանց համար ոչ ախտածին տեսակներով.
Burgey-ի ուղեցույցի 15-րդ և 16-րդ խմբերը միավորում են սահող բակտերիաները, որոնք պտղաբեր մարմիններ չեն կազմում և ձևավորում դրանք: Խմբերը չեն ներառում մարդկանց համար ախտածին տեսակներ:
Խումբ 17 Burgey-ի որոշիչ. Գրամ-դրական կոկի. Ներառում է Enterococcus Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus սեռերի պատեհապաշտ տեսակները։
Խումբ 18 Burgey-ի որոշիչ. Սպոր առաջացնող գրամ-դրական ձողեր և կոկիներ: Ներառում է Clostridium և Bacillus ցեղի պաթոգեն, պայմանականորեն ախտածին ձողեր։
Burgey-ի որոշիչի 19-րդ խումբ. Սպոր առաջացնող կանոնավոր ձևի գրամ դրական ձողեր: Ներառյալ Erysipelothrix և Listeria ցեղի պատեհապաշտ տեսակները։
Խումբ 20 Burgey-ի որոշիչ. Անկանոն ձևի սպոր ձևավորող գրամ-դրական ձողեր: Խմբում ընդգրկված են Actinomyces, Corynebacterium Gardnerella, Mobiluncus ցեղի ախտածին և պատեհապաշտ տեսակները։
Burgey-ի որոշիչի 21-րդ խումբ. Միկոբակտերիա. Ներառում է Mycobacterium միակ սեռը, որը միավորում է ախտածին և պատեհապաշտ տեսակները։
22-29 խմբեր. Ակտինոմիցետներ. Բազմաթիվ տեսակներից միայն նոկարդիոֆորմ ակտինոմիցետները (Խումբ 22)՝ Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi և Terrabacter սեռերի, որոնք կարող են ախտահարումներ առաջացնել մարդկանց մոտ:
Խումբ 30 Burgey-ի որոշիչ. Միկոպլազմաներ. Acholeplasma, Mycoplasma և Ureaplasma ցեղի մեջ ընդգրկված տեսակները ախտածին են մարդկանց համար:
Բերգեյի որոշիչի մնացած խմբերը՝ մեթանոգեն բակտերիաներ (31), սուլֆատ վերականգնող բակտերիաներ (32 ծայրահեղ հալոֆիլ աերոբ արխեեբակտերիա (33), բջջապատից զուրկ արխեբակտերիաներ (34), ծայրահեղ ջերմաֆիլներ և հիպերթերմոֆիլներ, ծծումբը մետաբոլիզացնող (35) պարունակում է մարդկանց համար պաթոգեն տեսակներ:
Բակտերիաները օրգանիզմների ամենահին խումբն են, որոնք ներկայումս գոյություն ունեն Երկրի վրա: Առաջին բակտերիաները, հավանաբար, հայտնվել են ավելի քան 3,5 միլիարդ տարի առաջ և գրեթե մեկ միլիարդ տարի եղել են մեր մոլորակի միակ կենդանի արարածները: Քանի որ սրանք վայրի բնության առաջին ներկայացուցիչներն էին, նրանց մարմինն ուներ պարզունակ կառուցվածք։
Ժամանակի ընթացքում դրանց կառուցվածքը դարձավ ավելի բարդ, սակայն նույնիսկ այսօր բակտերիաները համարվում են ամենապրիմիտիվ միաբջիջ օրգանիզմները։ Հետաքրքիր է, որ որոշ բակտերիաներ դեռ պահպանում են իրենց հին նախնիների պարզունակ առանձնահատկությունները: Սա նկատվում է բակտերիաների մոտ, որոնք ապրում են տաք ծծմբի աղբյուրներում և ջրամբարների հատակի անօքսիկ տիղմերում:
Բակտերիաների մեծ մասը անգույն է: Միայն մի քանիսն են գունավոր մանուշակագույն կամ կանաչ: Բայց շատ բակտերիաների գաղութները վառ գույն ունեն, ինչը պայմանավորված է շրջակա միջավայր գունավոր նյութի արտանետմամբ կամ բջիջների պիգմենտացիայով։
Բակտերիաների աշխարհի հայտնագործողը 17-րդ դարի հոլանդացի բնագետ Էնթոնի Լևենհուկն էր, ով առաջին անգամ ստեղծեց կատարյալ խոշորացույցի մանրադիտակ, որը մեծացնում է առարկաները 160-270 անգամ:
Բակտերիաները դասակարգվում են որպես պրոկարիոտներ և բաժանվում են առանձին թագավորության՝ բակտերիաների։
մարմնի ձեւը
Բակտերիաները բազմաթիվ և բազմազան օրգանիզմներ են։ Նրանք տարբերվում են ձևով.
| բակտերիաների անվանումը | Բակտերիաների ձևը | Բակտերիաների պատկեր |
| կոկկի | գնդաձեւ | |
| Բացիլուս |  | ձողաձև |
| Վիբրիո | կոր ստորակետ | |
| Spirillum |  | Պարույր |
| streptococci |  | Կոկիների շղթա |
| Ստաֆիլոկոկներ |  | Կոկիների կլաստերներ |
| դիպլոկոկներ | Երկու կլոր բակտերիաներ՝ փակված մեկ ցեխոտ պարկուճում |
Փոխադրման ուղիները
Բակտերիաների շարքում կան շարժական և անշարժ ձևեր։ Շարժականները շարժվում են ալիքանման կծկումների միջոցով կամ դրոշակների (ոլորված պտուտակավոր թելեր) օգնությամբ, որոնք կազմված են հատուկ ֆլագելինի սպիտակուցից։ Կարող է լինել մեկ կամ մի քանի դրոշակ: Դրանք որոշ բակտերիաներում տեղակայված են բջջի մի ծայրում, մյուսներում՝ երկու կամ ամբողջ մակերեսի վրա:

Բայց շարժումը բնորոշ է նաև շատ այլ բակտերիաների, որոնք չունեն դրոշակ: Այսպիսով, արտաքինից լորձով պատված բակտերիաները ունակ են սահող շարժման։
Ջրի և հողի որոշ բակտերիաներ՝ առանց դրոշակների, ունեն գազային վակուոլներ ցիտոպլազմայում: Բջջում կարող է լինել 40-60 վակուոլ: Նրանցից յուրաքանչյուրը լցված է գազով (ենթադրաբար ազոտով): Վակուոլներում գազի քանակությունը կարգավորելով՝ ջրային բակտերիաները կարող են սուզվել ջրի սյունի մեջ կամ բարձրանալ դրա մակերես, մինչդեռ հողի բակտերիաները կարող են շարժվել հողի մազանոթներում։
Հաբիթաթ
Կազմակերպման պարզության և ոչ հավակնոտության շնորհիվ բակտերիաները լայնորեն տարածված են բնության մեջ: Բակտերիաները հանդիպում են ամենուր՝ նույնիսկ ամենամաքուր աղբյուրի ջրի մի կաթիլում, հողի հատիկներում, օդում, ժայռերի վրա, բևեռային ձյան մեջ, անապատի ավազների վրա, օվկիանոսի հատակին, մեծ խորություններից արդյունահանվող յուղի մեջ և նույնիսկ տաք: աղբյուրի ջուր՝ մոտ 80ºС ջերմաստիճանով։ Նրանք ապրում են բույսերի, մրգերի, տարբեր կենդանիների և մարդկանց մոտ՝ աղիքներում, բերանում, վերջույթներում և մարմնի մակերեսին։
Բակտերիաները ամենափոքր և ամենաբազմաթիվ կենդանի արարածներն են։ Փոքր չափերի պատճառով դրանք հեշտությամբ թափանցում են ցանկացած ճաքերի, ճեղքերի, ծակոտիների մեջ։ Շատ դիմացկուն և հարմարեցված գոյության տարբեր պայմաններին: Նրանք հանդուրժում են չորացումը, ծայրահեղ ցրտերը, տաքանալը մինչև 90ºС՝ չկորցնելով կենսունակությունը։
Երկրի վրա գործնականում չկա մի տեղ, որտեղ բակտերիաներ չհայտնաբերվեին, բայց տարբեր քանակությամբ։ Բակտերիաների կենսապայմանները բազմազան են. Նրանցից ոմանք օդի թթվածնի կարիք ունեն, մյուսները՝ չունեն և կարողանում են ապրել թթվածնազուրկ միջավայրում։
Օդում. բակտերիաները բարձրանում են մթնոլորտի վերին շերտ մինչև 30 կմ: եւ ավելին.
Հատկապես նրանցից շատերը հողում են: Մեկ գրամ հողը կարող է պարունակել հարյուր միլիոնավոր բակտերիաներ։
Ջրի մեջ՝ բաց ջրամբարների մակերևութային ջրերի շերտերում։ Օգտակար ջրային բակտերիաները հանքայնացնում են օրգանական մնացորդները:
Կենդանի օրգանիզմներում՝ պաթոգեն բակտերիաները օրգանիզմ են ներթափանցում արտաքին միջավայրից, բայց միայն բարենպաստ պայմաններում առաջացնում են հիվանդություններ։ Սիմբիոտիկները ապրում են մարսողական օրգաններում՝ նպաստելով սննդի քայքայմանը և յուրացմանը, վիտամինների սինթեզմանը։
Արտաքին կառուցվածքը
Բակտերիաների բջիջը հագցված է հատուկ խիտ պատի մեջ՝ բջջային պատի մեջ, որն իրականացնում է պաշտպանիչ և օժանդակ գործառույթներ, ինչպես նաև տալիս է բակտերիային մշտական, բնորոշ ձև: Բակտերիաների բջջային պատը նման է բուսական բջիջի պատյանին։ Այն թափանցելի է. դրա միջոցով սննդանյութերն ազատորեն անցնում են բջիջ, իսկ նյութափոխանակության արտադրանքները դուրս են գալիս շրջակա միջավայր: Բակտերիաները հաճախ զարգացնում են լորձի լրացուցիչ պաշտպանիչ շերտ՝ պարկուճ, բջջային պատի վրա։ Պարկուճի հաստությունը կարող է շատ անգամ ավելի մեծ լինել, քան բջջի տրամագիծը, բայց այն կարող է լինել շատ փոքր: Պարկուճը բջջի պարտադիր մաս չէ, այն ձևավորվում է կախված բակտերիաների ներթափանցման պայմաններից։ Այն կանխում է բակտերիաների չորացումը:
Որոշ բակտերիաների մակերեսին կան երկար դրոշակներ (մեկ, երկու կամ շատ) կամ կարճ բարակ վիլլիներ։ Դրոշակի երկարությունը կարող է մի քանի անգամ ավելի մեծ լինել, քան բակտերիայի մարմնի չափը։ Բակտերիաները շարժվում են դրոշակների և վիլլի օգնությամբ։

Ներքին կառուցվածքը
Բակտերիալ բջջի ներսում խիտ անշարժ ցիտոպլազմա է: Այն ունի շերտավոր կառուցվածք, չկա վակուոլներ, ուստի տարբեր սպիտակուցներ (ֆերմենտներ) և պահուստային սննդանյութեր տեղակայված են հենց ցիտոպլազմայի նյութում։ Բակտերիալ բջիջները կորիզ չունեն։ Նրանց բջիջների կենտրոնական մասում խտացված է ժառանգական տեղեկատվություն կրող նյութ։ Բակտերիաներ, - նուկլեինաթթու - ԴՆԹ: Բայց այս նյութը միջուկում շրջանակված չէ:
Բակտերիալ բջջի ներքին կազմակերպումը բարդ է և ունի իր առանձնահատկությունները: Բջջային պատից ցիտոպլազմը բաժանվում է ցիտոպլազմային թաղանթով։ Ցիտոպլազմայում առանձնանում են հիմնական նյութը կամ մատրիցը, ռիբոսոմները և մի շարք գործառույթներ կատարող թաղանթային կառուցվածքների փոքր քանակություն (միտոքոնդրիաների, էնդոպլազմային ցանցի, Գոլջիի ապարատի անալոգներ): Բակտերիալ բջիջների ցիտոպլազմը հաճախ պարունակում է տարբեր ձևերի և չափերի հատիկներ: Հատիկները կարող են կազմված լինել միացություններից, որոնք ծառայում են որպես էներգիայի և ածխածնի աղբյուր: Բակտերիալ բջջում հայտնաբերվում են նաև ճարպի կաթիլներ։
Բջջի կենտրոնական մասում միջուկային նյութը՝ ԴՆԹ-ն, տեղայնացված է, ցիտոպլազմայից թաղանթով առանձնացված չէ։ Սա միջուկի անալոգն է՝ նուկլեոիդը։ Նուկլեոիդը չունի թաղանթ, միջուկ և քրոմոսոմների հավաքածու։
Սնուցման մեթոդներ
Բակտերիաները կերակրման տարբեր եղանակներ ունեն։ Դրանցից են ավտոտրոֆներն ու հետերոտրոֆները։ Ավտոտրոֆները օրգանիզմներ են, որոնք կարող են ինքնուրույն ձևավորել օրգանական նյութեր իրենց սնուցման համար:
Բույսերը ազոտի կարիք ունեն, բայց նրանք իրենք չեն կարող օդից ազոտ կլանել։ Որոշ բակտերիաներ օդում ազոտի մոլեկուլները միացնում են այլ մոլեկուլների հետ, ինչի արդյունքում բույսերին հասանելի են նյութեր:
Այս բակտերիաները տեղավորվում են երիտասարդ արմատների բջիջներում, ինչը հանգեցնում է արմատների վրա խտացումների առաջացմանը, որոնք կոչվում են հանգույցներ: Նման հանգույցներ առաջանում են լոբազգիների ընտանիքի բույսերի և որոշ այլ բույսերի արմատների վրա։

Արմատները բակտերիաներին ապահովում են ածխաջրերով, իսկ բակտերիաները արմատներին տալիս են ազոտ պարունակող նյութեր, որոնք կարող են կլանել բույսը։ Նրանց հարաբերությունները փոխշահավետ են։
Բույսերի արմատները արտազատում են բազմաթիվ օրգանական նյութեր (շաքարներ, ամինաթթուներ և այլն), որոնցով սնվում են բակտերիաները։ Հետեւաբար, հատկապես շատ բակտերիաներ նստում են արմատները շրջապատող հողի շերտում։ Այս բակտերիաները մահացած բույսերի մնացորդները վերածում են բույսին հասանելի նյութերի: Հողի այս շերտը կոչվում է ռիզոսֆերա։
Կան մի քանի վարկածներ հանգուցային բակտերիաների արմատային հյուսվածքների ներթափանցման վերաբերյալ.
- էպիդերմիսի և կեղևի հյուսվածքի վնասման միջոցով;
- արմատային մազերի միջոցով;
- միայն երիտասարդ բջջային թաղանթի միջոցով;
- պեկտինոլիտիկ ֆերմենտներ արտադրող ուղեկից բակտերիաների պատճառով;
- տրիպտոֆանից B-ինդոլաքացախաթթվի սինթեզի խթանման շնորհիվ, որը միշտ առկա է բույսերի արմատային սեկրեցներում:
Հանգույց բակտերիաների արմատային հյուսվածքի մեջ ներմուծման գործընթացը բաղկացած է երկու փուլից.
- արմատային մազերի վարակ;
- հանգույցների ձևավորման գործընթացը.
Շատ դեպքերում ներխուժող բջիջը ակտիվորեն բազմանում է, ձևավորում, այսպես կոչված, վարակիչ թելեր և արդեն նման թելերի տեսքով տեղափոխվում է բույսերի հյուսվածքներ։ Հանգույց բակտերիաները, որոնք առաջացել են վարակիչ թելից, շարունակում են բազմանալ հյուրընկալող հյուսվածքում:

Լցված հանգուցային բակտերիաների արագ բազմացող բջիջներով՝ բույսերի բջիջները սկսում են ինտենսիվ բաժանվել։ Երիտասարդ հանգույցի կապը հատիկավոր բույսի արմատի հետ իրականացվում է անոթային-թելքավոր կապոցների շնորհիվ։ Գործելու ժամանակահատվածում հանգույցները սովորաբար խիտ են լինում։ Օպտիմալ ակտիվության դրսևորման պահին հանգույցները ձեռք են բերում վարդագույն գույն (լեգոգլոբինի պիգմենտի շնորհիվ)։ Միայն այն բակտերիաները, որոնք պարունակում են լեգոգլոբին, կարող են ամրացնել ազոտը։
Հանգույց բակտերիաները ստեղծում են տասնյակ և հարյուրավոր կիլոգրամ ազոտական պարարտանյութեր մեկ հեկտար հողի վրա:
Նյութափոխանակություն
Բակտերիաները միմյանցից տարբերվում են նյութափոխանակության մեջ։ Ոմանց մոտ այն անցնում է թթվածնի մասնակցությամբ, մյուսների մոտ՝ առանց նրա մասնակցության։
Բակտերիաների մեծ մասը սնվում է պատրաստի օրգանական նյութերով։ Դրանցից միայն մի քանիսը (կապտա-կանաչ, կամ ցիանոբակտերիաներ) կարող են օրգանական նյութեր ստեղծել անօրգանականներից։ Նրանք կարևոր դեր են խաղացել Երկրի մթնոլորտում թթվածնի կուտակման գործում։
Բակտերիաները դրսից կլանում են նյութերը, պոկում են դրանց մոլեկուլները, հավաքում իրենց պատյանը այս մասերից և լրացնում դրանց պարունակությունը (այսպես են աճում) և դուրս են նետում ավելորդ մոլեկուլները։ Մանրէի պատյանը և թաղանթը թույլ են տալիս նրան կլանել միայն ճիշտ նյութերը։

Եթե մանրէի թաղանթն ու թաղանթը լիովին անթափանց լինեին, ոչ մի նյութ բջիջ չէր մտնի։ Եթե դրանք թափանցելի լինեին բոլոր նյութերի համար, ապա բջջի պարունակությունը կխառնվեր միջավայրի հետ՝ այն լուծույթին, որում ապրում է բակտերիան: Բակտերիաների գոյատևման համար անհրաժեշտ է պատյան, որը թույլ է տալիս անցնել անհրաժեշտ նյութերը, բայց ոչ նրանք, որոնք անհրաժեշտ չեն:
Բակտերիան կլանում է իր մոտ գտնվող սնուցիչները։ Ի՞նչ կլինի հետո։ Եթե այն կարող է ինքնուրույն շարժվել (դրոշակը շարժելով կամ լորձը հետ մղելով), ապա այն շարժվում է այնքան ժամանակ, մինչև գտնի անհրաժեշտ նյութերը։

Եթե այն չի կարող շարժվել, ապա սպասում է, մինչև դիֆուզիան (մի նյութի մոլեկուլների՝ մեկ այլ նյութի մոլեկուլների հաստությունը ներթափանցելու ունակությունը) բերի նրան անհրաժեշտ մոլեկուլները։
Բակտերիաները միկրոօրգանիզմների այլ խմբերի հետ միասին հսկայական քիմիական աշխատանք են կատարում։ Փոխակերպելով տարբեր միացություններ՝ նրանք ստանում են իրենց կենսագործունեության համար անհրաժեշտ էներգիան և սննդանյութերը։ Բազմազան են նյութափոխանակության պրոցեսները, էներգիա ստանալու ուղիները և դրանց մարմնի նյութերը բակտերիաներում կառուցվող նյութերի անհրաժեշտությունը։
Այլ բակտերիաները բավարարում են ածխածնի բոլոր կարիքները, որոնք անհրաժեշտ են մարմնի օրգանական նյութերի սինթեզի համար անօրգանական միացությունների հաշվին։ Նրանք կոչվում են ավտոտրոֆներ: Ավտոտրոֆ բակտերիաները կարողանում են օրգանական նյութեր սինթեզել անօրգանականներից։ Դրանց թվում առանձնանում են.
Քիմոսինթեզ
Ճառագայթային էներգիայի օգտագործումը ածխաթթու գազից և ջրից օրգանական նյութեր ստեղծելու ամենակարևոր, բայց ոչ միակ միջոցն է: Հայտնի են բակտերիաներ, որոնք օգտագործում են ոչ թե արևի լույսը որպես էներգիայի աղբյուր նման սինթեզի համար, այլ օրգանիզմների բջիջներում առաջացող քիմիական կապերի էներգիան որոշ անօրգանական միացությունների՝ ջրածնի սուլֆիդ, ծծումբ, ամոնիակ, ջրածին, ազոտական թթու, գունավոր միացությունների օքսիդացման ժամանակ։ երկաթ և մանգան: Նրանք օգտագործում են օրգանական նյութերը, որոնք առաջացել են այս քիմիական էներգիայի միջոցով՝ իրենց մարմնի բջիջները կառուցելու համար: Հետեւաբար, այս գործընթացը կոչվում է քիմոսինթեզ:
Քիմոսինթետիկ միկրոօրգանիզմների ամենակարևոր խումբը նիտրացնող բակտերիաներն են։ Այս բակտերիաները ապրում են հողում և իրականացնում են օրգանական մնացորդների քայքայման ժամանակ առաջացած ամոնիակի օքսիդացումը մինչև ազոտաթթու։ Վերջինս, արձագանքելով հողի հանքային միացությունների հետ, վերածվում է ազոտաթթվի աղերի։ Այս գործընթացը տեղի է ունենում երկու փուլով.
Երկաթի բակտերիաները սեւ երկաթը վերածում են օքսիդի: Առաջացած երկաթի հիդրօքսիդը նստում է և ձևավորում այսպես կոչված ճահճային երկաթի հանքաքար։
Որոշ միկրոօրգանիզմներ գոյություն ունեն մոլեկուլային ջրածնի օքսիդացման շնորհիվ՝ դրանով իսկ ապահովելով սնվելու ավտոտրոֆ եղանակ։
Ջրածնային բակտերիաների բնորոշ հատկանիշը հետերոտրոֆ ապրելակերպի անցնելու ունակությունն է, երբ այն ապահովված է օրգանական միացություններով և ջրածնի բացակայության դեպքում:
Այսպիսով, քիմոավտոտրոֆները բնորոշ ավտոտրոֆներ են, քանի որ նրանք ինքնուրույն սինթեզում են անհրաժեշտ օրգանական միացությունները անօրգանական նյութերից և դրանք պատրաստ չեն վերցնում այլ օրգանիզմներից, ինչպիսիք են հետերոտրոֆները: Քիմիաավտոտրոֆ բակտերիաները տարբերվում են ֆոտոտրոֆ բույսերից՝ որպես էներգիայի աղբյուր լույսից իրենց լիակատար անկախությամբ։
բակտերիալ ֆոտոսինթեզ
Որոշ պիգմենտ պարունակող ծծմբային բակտերիաներ (մանուշակագույն, կանաչ), որոնք պարունակում են հատուկ պիգմենտներ՝ բակտերիոքլորոֆիլներ, կարողանում են կլանել արևի էներգիան, որի օգնությամբ ջրածնի սուլֆիդը տրոհվում է նրանց օրգանիզմներում և տալիս ջրածնի ատոմներ՝ վերականգնելու համապատասխան միացությունները։ Այս գործընթացը շատ ընդհանրություններ ունի ֆոտոսինթեզի հետ և տարբերվում է միայն նրանով, որ մանուշակագույն և կանաչ բակտերիաներում ջրածնի սուլֆիդը (երբեմն կարբոքսիլաթթուներ) ջրածնի դոնոր է, իսկ կանաչ բույսերում՝ ջուր։ Դրանցում և մյուսներում ջրածնի պառակտումն ու փոխանցումն իրականացվում է կլանված արևի ճառագայթների էներգիայի շնորհիվ։
Նման բակտերիալ ֆոտոսինթեզը, որը տեղի է ունենում առանց թթվածնի արտազատման, կոչվում է ֆոտոռեդուկցիա։ Ածխածնի երկօքսիդի ֆոտովերականգնումը կապված է ջրածնի փոխանցման հետ ոչ թե ջրից, այլ ջրածնի սուլֆիդից.
6CO 2 + 12H 2 S + hv → C6H 12 O 6 + 12S \u003d 6H 2 O
Քիմոսինթեզի և բակտերիալ ֆոտոսինթեզի կենսաբանական նշանակությունը մոլորակային մասշտաբով համեմատաբար փոքր է։ Բնության մեջ ծծմբի ցիկլում էական դեր են խաղում միայն քիմոսինթետիկ բակտերիաները։ Ծծմբաթթվի աղերի տեսքով կանաչ բույսերի կողմից կլանված ծծումբը վերականգնվում է և դառնում սպիտակուցի մոլեկուլների մաս: Այնուհետև, փտած բակտերիաների կողմից մահացած բույսերի և կենդանիների մնացորդների ոչնչացման ժամանակ ծծումբն արտազատվում է ջրածնի սուլֆիդի տեսքով, որը ծծմբի բակտերիայով օքսիդանում է մինչև ազատ ծծումբ (կամ ծծմբաթթու), որը կազմում է հողում բույսերի համար հասանելի սուլֆիտներ: Քիմիա և ֆոտոավտոտրոֆ բակտերիաները կարևոր են ազոտի և ծծմբի ցիկլում:
սպորացում
Սպորները ձևավորվում են բակտերիաների բջջի ներսում: Սպորի ձևավորման գործընթացում բակտերիալ բջիջը ենթարկվում է մի շարք կենսաքիմիական գործընթացների: Դրանում նվազում է ազատ ջրի քանակը, նվազում է ֆերմենտային ակտիվությունը։ Սա ապահովում է սպորների դիմադրությունը շրջակա միջավայրի անբարենպաստ պայմաններին (բարձր ջերմաստիճան, աղի բարձր կոնցենտրացիա, չորացում և այլն): Սպորների առաջացումը բնորոշ է մանրէների միայն փոքր խմբին։

Սպորները բակտերիաների կյանքի ցիկլի էական փուլ չեն: Սպորացումը սկսվում է միայն սննդանյութերի պակասից կամ նյութափոխանակության արտադրանքի կուտակումից: Սպորների տեսքով բակտերիաները կարող են երկար ժամանակ քնած մնալ։ Բակտերիալ սպորները դիմանում են երկարատև եռման և շատ երկար սառեցման: Երբ բարենպաստ պայմաններ են առաջանում, վեճը բողբոջում է և դառնում կենսունակ։ Բակտերիալ սպորները հարմարեցված են անբարենպաստ պայմաններում գոյատևելու համար:
վերարտադրություն
Բակտերիաները բազմանում են՝ մեկ բջիջը երկուսի բաժանելով։ Որոշակի չափի հասնելով՝ բակտերիան բաժանվում է երկու միանման բակտերիաների։ Հետո նրանցից յուրաքանչյուրը սկսում է կերակրել, աճել, բաժանվել և այլն։

Բջջի երկարացումից հետո աստիճանաբար ձևավորվում է լայնակի միջնապատ, այնուհետև դուստր բջիջները շեղվում են. Բազմաթիվ բակտերիաներում, որոշակի պայմաններում, բջիջները բաժանումից հետո մնում են միացված բնորոշ խմբերով: Այս դեպքում, կախված բաժանման հարթության ուղղությունից և բաժանումների քանակից, առաջանում են տարբեր ձևեր։ Բացառության կարգով վերարտադրումը բողբոջումներով տեղի է ունենում բակտերիաների մոտ:
Բարենպաստ պայմաններում բազմաթիվ բակտերիաների բջիջների բաժանումը տեղի է ունենում 20-30 րոպեն մեկ: Նման արագ բազմացման դեպքում մեկ բակտերիաի սերունդը 5 օրվա ընթացքում կարողանում է զանգված կազմել, որը կարող է լցնել բոլոր ծովերն ու օվկիանոսները։ Պարզ հաշվարկը ցույց է տալիս, որ օրական կարող է ձևավորվել 72 սերունդ (720,000,000,000,000,000,000 բջիջ): Եթե թարգմանվի քաշի` 4720 տոննա: Այնուամենայնիվ, բնության մեջ դա տեղի չի ունենում, քանի որ բակտերիաների մեծ մասը արագ մահանում է արևի լույսի, չորացման, սննդի պակասի, 65-100ºС տաքացման, տեսակների միջև պայքարի և այլնի ազդեցության տակ:
Բակտերիան (1), կլանելով բավականաչափ սնունդ, մեծանում է չափսերով (2) և սկսում պատրաստվել վերարտադրության (բջիջների բաժանման): Նրա ԴՆԹ-ն (մանրէի մեջ ԴՆԹ-ի մոլեկուլը փակված է օղակով) կրկնապատկվում է (մանրէը արտադրում է այս մոլեկուլի կրկնօրինակը)։ ԴՆԹ-ի երկու մոլեկուլներն էլ (3.4) կարծես կցված են բակտերիաների պատին և, երբ երկարացվում են, բակտերիաները շեղվում են կողքերից (5.6): Նախ, նուկլեոտիդը բաժանվում է, ապա ցիտոպլազմա:
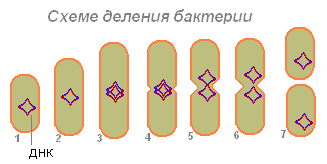
Բակտերիաների վրա ԴՆԹ-ի երկու մոլեկուլների շեղումից հետո առաջանում է կծկում, որն աստիճանաբար բակտերիայի մարմինը բաժանում է երկու մասի, որոնցից յուրաքանչյուրը պարունակում է ԴՆԹ մոլեկուլ (7)։
Դա տեղի է ունենում (խոտի բացիլում), երկու բակտերիաներ կպչում են միմյանց, և նրանց միջև կամուրջ է առաջանում (1,2):

ԴՆԹ-ն տեղափոխվում է մի բակտերիայից մյուսը jumper-ի միջոցով (3): Մեկ բակտերիայից մեկում ԴՆԹ-ի մոլեկուլները միահյուսվում են, որոշ տեղերում կպչում իրար (4), որից հետո փոխանակում են հատվածները (5):
Բակտերիաների դերը բնության մեջ
Շրջանառություն
Բակտերիաները բնության մեջ նյութերի ընդհանուր շրջանառության ամենակարեւոր օղակն են։ Բույսերը բարդ օրգանական նյութեր են ստեղծում ածխաթթու գազից, ջրից և հողի հանքային աղերից։ Այս նյութերը հող են վերադառնում սատկած սնկերի, բույսերի և կենդանիների դիակներով։ Բակտերիաները բարդ նյութերը քայքայում են պարզ նյութերի, որոնք վերաօգտագործվում են բույսերի կողմից։
Բակտերիաները ոչնչացնում են մահացած բույսերի և կենդանիների դիակների բարդ օրգանական նյութերը, կենդանի օրգանիզմների արտազատումները և տարբեր թափոնները: Սնվելով այս օրգանական նյութերով՝ սապրոֆիտ քայքայվող բակտերիաները դրանք վերածում են հումուսի։ Սրանք մեր մոլորակի կարգապահներն են: Այսպիսով, բակտերիաները ակտիվորեն ներգրավված են բնության նյութերի ցիկլում:
հողի ձևավորում
Քանի որ բակտերիաները տարածված են գրեթե ամենուր և հայտնաբերված հսկայական քանակությամբ, նրանք մեծապես որոշում են բնության մեջ տեղի ունեցող տարբեր գործընթացները: Աշնանը թափվում են ծառերի ու թփերի տերեւները, սատկում են վերգետնյա խոտի ընձյուղները, թափվում են հին ճյուղերը, ժամանակ առ ժամանակ թափվում են ծեր ծառերի բները։ Այս ամենն աստիճանաբար վերածվում է հումուսի։ 1 սմ 3-ում: Անտառային հողի մակերևութային շերտը պարունակում է մի քանի տեսակների հարյուր միլիոնավոր հողի սապրոֆիտ բակտերիաներ։ Այս բակտերիաները հումուսը վերածում են տարբեր հանքանյութերի, որոնք կարող են ներծծվել հողից բույսերի արմատներով:
Հողի որոշ բակտերիաներ ունակ են կլանել օդից ազոտը՝ այն օգտագործելով կյանքի գործընթացներում։ Ազոտը ամրագրող այս բակտերիաները ապրում են ինքնուրույն կամ բնակվում են հատիկավոր բույսերի արմատներում: Ներթափանցելով հատիկաընդեղենների արմատները՝ այս բակտերիաները առաջացնում են արմատային բջիջների աճ և դրանց վրա հանգույցների ձևավորում։
Այս բակտերիաները արձակում են ազոտային միացություններ, որոնք օգտագործում են բույսերը: Բակտերիաները բույսերից ստանում են ածխաջրեր և հանքային աղեր։ Այսպիսով, սերտ կապ կա հատիկավոր բույսի և հանգուցային բակտերիաների միջև, ինչը օգտակար է և՛ մեկի, և՛ մյուս օրգանիզմի համար։ Այս երեւույթը կոչվում է սիմբիոզ։
Հանգույցային բակտերիաների հետ իրենց սիմբիոզի շնորհիվ հատիկաբույսերը հարստացնում են հողը ազոտով՝ նպաստելով բերքատվության ավելացմանը։
Բաշխումը բնության մեջ
Միկրոօրգանիզմները ամենուր են: Բացառություն են կազմում ակտիվ հրաբուխների խառնարանները և պայթեցված ատոմային ռումբերի էպիկենտրոններում գտնվող փոքր տարածքները: Ո՛չ Անտարկտիկայի ցածր ջերմաստիճանը, ո՛չ գեյզերների եռացող շիթերը, ո՛չ աղի ավազաններում հագեցած աղի լուծույթները, ո՛չ լեռների գագաթների ուժեղ մեկուսացումը, ո՛չ միջուկային ռեակտորների խիստ ճառագայթումը չեն խանգարում միկրոֆլորայի գոյությանը և զարգացմանը: Բոլոր կենդանի էակները մշտապես փոխազդում են միկրոօրգանիզմների հետ՝ հաճախ լինելով ոչ միայն դրանց պահեստները, այլև տարածողները: Միկրոօրգանիզմները մեր մոլորակի բնիկներն են, որոնք ակտիվորեն զարգացնում են ամենաանհավանական բնական սուբստրատները:
Հողի միկրոֆլորա
Հողի մեջ բակտերիաների թիվը չափազանց մեծ է՝ հարյուր միլիոնավոր և միլիարդավոր անհատներ 1 գրամում: Նրանք շատ ավելի առատ են հողում, քան ջրում և օդում։ Հողերում բակտերիաների ընդհանուր թիվը տարբեր է։ Բակտերիաների թիվը կախված է հողի տեսակից, նրանց վիճակից, շերտերի խորությունից։
Հողի մասնիկների մակերեսին միկրոօրգանիզմները տեղակայված են փոքր միկրոգաղութներում (յուրաքանչյուրը 20-100 բջիջ): Հաճախ դրանք զարգանում են օրգանական նյութերի խցանումների հաստությամբ, կենդանի և մեռնող բույսերի արմատների վրա, բարակ մազանոթներում և ներսի գնդիկներով։
Հողի միկրոֆլորան շատ բազմազան է. Այստեղ հանդիպում են բակտերիաների ֆիզիոլոգիական տարբեր խմբեր՝ փտած, նիտրացնող, ազոտամիկացնող, ծծմբային բակտերիաներ և այլն։ Միկրոֆլորան հողի գոյացման գործոններից է։
Հողի մեջ միկրոօրգանիզմների զարգացման տարածքը կենդանի բույսերի արմատներին հարող գոտին է: Այն կոչվում է ռիզոսֆերա, իսկ դրա մեջ պարունակվող միկրոօրգանիզմների ամբողջությունը կոչվում է ռիզոսֆերային միկրոֆլորա։
Ջրամբարների միկրոֆլորան
Ջուրը բնական միջավայր է, որտեղ մեծ քանակությամբ միկրոօրգանիզմներ են աճում: Նրանց մեծ մասը ջուրը մտնում է հողից։ Գործոն, որը որոշում է ջրի մեջ բակտերիաների քանակը, դրա մեջ սննդանյութերի առկայությունը։ Ամենամաքուրը արտեզյան հորերի և աղբյուրների ջրերն են։ Բաց ջրամբարներն ու գետերը շատ հարուստ են մանրէներով։ Բակտերիաների ամենամեծ քանակությունը հայտնաբերված է ջրի մակերեսային շերտերում՝ ափին ավելի մոտ։ Ափից հեռավորության և խորության ավելացման հետ բակտերիաների թիվը նվազում է։
Մաքուր ջուրը պարունակում է 100-200 բակտերիաներ 1 մլ-ում, մինչդեռ աղտոտված ջուրը պարունակում է 100-300 հազար և ավելի: Ներքևի տիղմում կան բազմաթիվ բակտերիաներ, հատկապես մակերեսային շերտում, որտեղ մանրէները թաղանթ են կազմում։ Այս թաղանթում կան շատ ծծմբի և երկաթի բակտերիաներ, որոնք օքսիդացնում են ջրածնի սուլֆիդը մինչև ծծմբաթթու և դրանով իսկ կանխում ձկների մահը: Տիղմում ավելի շատ սպորակիր ձևեր կան, իսկ ջրում գերակշռում են ոչ սպորաբեր ձևերը։
Տեսակային կազմով ջրային միկրոֆլորան նման է հողի միկրոֆլորային, սակայն հանդիպում են նաև հատուկ ձևեր։ Ոչնչացնելով ջուրն ընկած տարատեսակ թափոնները՝ միկրոօրգանիզմները աստիճանաբար իրականացնում են ջրի այսպես կոչված կենսաբանական մաքրումը։
Օդի միկրոֆլորա
Օդի միկրոֆլորան ավելի քիչ է, քան հողի և ջրի միկրոֆլորան: Բակտերիաները օդ են բարձրանում փոշու հետ, կարող են որոշ ժամանակ մնալ այնտեղ, այնուհետև նստել երկրի մակերեսին և մահանալ սննդի պակասից կամ ուլտրամանուշակագույն ճառագայթների ազդեցության տակ: Օդում միկրոօրգանիզմների թիվը կախված է աշխարհագրական տարածքից, տեղանքից, սեզոնից, փոշու աղտոտվածությունից և այլն: Փոշու յուրաքանչյուր բծը միկրոօրգանիզմների կրող է: Բակտերիաների մեծ մասը օդում է արդյունաբերական ձեռնարկությունների վրա: Գյուղի օդն ավելի մաքուր է։ Ամենամաքուր օդը անտառների, լեռների, ձնառատ տարածքների վրա է։ Օդի վերին շերտերն ավելի քիչ մանրէներ են պարունակում։ Օդի միկրոֆլորայում կան բազմաթիվ պիգմենտային և սպորակիր բակտերիաներ, որոնք մյուսներից ավելի դիմացկուն են ուլտրամանուշակագույն ճառագայթների նկատմամբ:
Մարդու մարմնի միկրոֆլորան
Մարդու մարմինը, նույնիսկ լիովին առողջ, միշտ միկրոֆլորայի կրող է։ Երբ մարդու մարմինը շփվում է օդի և հողի հետ, մի շարք միկրոօրգանիզմներ, այդ թվում՝ հարուցիչներ (տետանուսի բացիլներ, գազային գանգրենա և այլն), նստում են հագուստի և մաշկի վրա։ Մարդու մարմնի բաց մասերը ամենից հաճախ աղտոտված են: Ձեռքերին հայտնաբերվում են E. coli, staphylococci: Բերանի խոռոչում կան ավելի քան 100 տեսակի մանրէներ։ Բերանը իր ջերմաստիճանով, խոնավությամբ, սննդանյութերի մնացորդներով հիանալի միջավայր է միկրոօրգանիզմների զարգացման համար։
Ստամոքսը թթվային ռեակցիա է ունենում, ուստի դրա մեջ միկրոօրգանիզմների մեծ մասը մահանում է: Բարակ աղիքից սկսած՝ ռեակցիան դառնում է ալկալային, այսինքն. բարենպաստ մանրէների համար. Հաստ աղիքի միկրոֆլորան շատ բազմազան է։ Յուրաքանչյուր մեծահասակ օրական արտազատում է մոտ 18 միլիարդ բակտերիա արտաթորանքով, այսինքն. ավելի շատ անհատներ, քան մարդիկ ամբողջ աշխարհում:
Ներքին օրգանները, որոնք կապված չեն արտաքին միջավայրի հետ (ուղեղ, սիրտ, լյարդ, միզապարկ և այլն), սովորաբար զերծ են մանրէներից։ Մանրէները այս օրգաններ են մտնում միայն հիվանդության ժամանակ։
Բակտերիաներ հեծանվավազքի մեջ
Միկրոօրգանիզմներն ընդհանրապես և մանրէները հատկապես կարևոր դեր են խաղում Երկրի վրա նյութի կենսաբանորեն կարևոր ցիկլերում՝ իրականացնելով քիմիական փոխակերպումներ, որոնք լիովին անհասանելի են ոչ բույսերի, ոչ կենդանիների համար: Տարրերի ցիկլի տարբեր փուլեր իրականացվում են տարբեր տեսակի օրգանիզմների կողմից։ Օրգանիզմների յուրաքանչյուր առանձին խմբի գոյությունը կախված է այլ խմբերի կողմից իրականացվող տարրերի քիմիական փոխակերպումից։
ազոտի ցիկլը
Ազոտային միացությունների ցիկլային փոխակերպումը կարևոր դեր է խաղում կենսոլորտային տարբեր օրգանիզմներին ազոտի անհրաժեշտ ձևերի մատակարարման հարցում՝ սննդային կարիքների տեսանկյունից: Ընդհանուր ազոտի ֆիքսացիայի ավելի քան 90%-ը պայմանավորված է որոշակի բակտերիաների նյութափոխանակության ակտիվությամբ:
Ածխածնի ցիկլը
Օրգանական ածխածնի կենսաբանական վերափոխումը ածխածնի երկօքսիդի, որն ուղեկցվում է մոլեկուլային թթվածնի կրճատմամբ, պահանջում է տարբեր միկրոօրգանիզմների համատեղ նյութափոխանակության ակտիվություն: Շատ աերոբ բակտերիաներ իրականացնում են օրգանական նյութերի ամբողջական օքսիդացում։ Աերոբիկ պայմաններում օրգանական միացությունները սկզբնապես քայքայվում են խմորումով, և ֆերմենտացման օրգանական վերջնական արտադրանքը հետագայում օքսիդացվում է անաէրոբ շնչառության միջոցով, եթե առկա են անօրգանական ջրածնի ընդունիչներ (նիտրատ, սուլֆատ կամ CO2):
Ծծմբի ցիկլը
Կենդանի օրգանիզմների համար ծծումբը հասանելի է հիմնականում լուծվող սուլֆատների կամ նվազեցված օրգանական ծծմբի միացությունների տեսքով։
Երկաթի ցիկլը
Որոշ քաղցրահամ ջրի ջրամբարներ պարունակում են երկաթի նվազեցված աղերի բարձր կոնցենտրացիաներ: Նման վայրերում զարգանում է կոնկրետ բակտերիալ միկրոֆլորա՝ երկաթի բակտերիաներ, որոնք օքսիդացնում են կրճատված երկաթը։ Նրանք մասնակցում են ճահճային երկաթի հանքաքարերի և երկաթի աղերով հարուստ ջրի աղբյուրների ձևավորմանը։
Բակտերիաները ամենահին օրգանիզմներն են, որոնք հայտնվել են մոտ 3,5 միլիարդ տարի առաջ Արքեում: Մոտ 2,5 միլիարդ տարի նրանք գերիշխում էին Երկրի վրա՝ ձևավորելով կենսոլորտը և մասնակցել թթվածնային մթնոլորտի ձևավորմանը։
Բակտերիաները ամենապարզ դասավորված կենդանի օրգանիզմներից են (բացառությամբ վիրուսների)։ Ենթադրվում է, որ դրանք Երկրի վրա հայտնված առաջին օրգանիզմներն են:
բակտերիաներ- Սրանք միաբջիջ, քլորոֆիլազուրկ օրգանիզմներ են, որոնք վեգետատիվ բազմանում են բաժանման, ավելի քիչ հաճախ ժանյակավոր, երբեմն ներբջջային սպորներ առաջացնելով։
Բակտերիաների չափերը չափվում են միկրոններով և, հազվադեպ բացառություններով, տատանվում են 0,06-0,3-ից մինչև 3-5 μ: Ջրի մեկ կաթիլը հեշտությամբ կարող է տեղավորել մի քանի հարյուր միլիոն բակտերիաներ:
Բակտերիալ բջջի ձևը բավականին միատեսակ է։ Հայտնի են բակտերիաների երեք հիմնական ձևեր՝ կլոր, ձողաձև և ոլորված՝ դրանց միջև բազմաթիվ և աննկատ անցումներով: Անտոն Դե Բարին դրանք պատկերավոր կերպով համեմատեց բիլիարդի գնդակի, մատիտի և խցանահանի հետ։
Կլոր ձևով բակտերիաները կոչվում են կոկկի: Նրանք տարբերվում են չափերով և հարաբերական դիրքով։ Զույգ-զույգ միացված կոկիկները կոչվում են դիպլոկոկներ, իսկ վզնոցի տեսքով միացվածները կոչվում են streptococci։ Բաժանվելիս երկու փոխադարձ ուղղահայաց ուղղություններով հերթափոխով առաջանում են տետրակոկներ։ Եթե բաժանումը ճիշտ է և կրկնվում է երեք փոխադարձ ուղղահայաց ուղղություններով, ապա բջջային կապերը ձևավորվում են փաթեթների տեսքով՝ դրանք այսպես կոչված սարդինաներն են։ Առանց մեծ օրինաչափության բաժանվելով տարբեր ուղղություններով՝ կոկիկները ձևավորում են պատահական ողկույզներ, որոնք հիշեցնում են խաղողի խոզանակ: Նրանք կոչվում են ստաֆիլոկոկներ:
Ձողաձև բակտերիաներն իրենց արտաքին տեսքով մի փոքր ավելի բազմազան են։ Դրանք կարող են լինել կտրված կամ կլորացված ծայրերով, գլանաձև, տակառաձև կամ կիտրոնի ձևով և, ասես, մեջտեղում սեղմված, հաճախ էլիպսաձև, որոնք տարբերվում են միայն լայնությամբ և երկարությամբ։ Ձողիկները կարող են լինել ուղիղ, կոր, միայնակ, զույգերով կամ շղթայով կապված, կարճ կամ խիստ երկարաձգված: Ձողաձև բակտերիաները, որոնց երկարությունը երկու անգամ կամ ավելի է լայնությունից, կոչվում են բացիլներ; եթե երկարության և լայնության տարբերությունը փոքր է, ապա դրանք կոչվում են բակտերիաներ:
Գանգուր մանրէները տարբերվում են ոչ միայն երկարությամբ ու հաստությամբ, այլեւ գանգուրների քանակով ու բնույթով։ Թեթևակի կորացած բակտերիաները (գանգուրը չի գերազանցում պարույրի 1/4 պտույտը) կոչվում են վիբրիո, մեկ կամ մի քանի խոշոր կանոնավոր գանգուրներով բակտերիաները կոչվում են սպիրիլլա: Խճճված ձևի երկար և բարակ բակտերիաները բազմաթիվ փոքր գանգուրներով, երբեմն ամբողջ թելիկի մեծ կորություններով, կոչվում են սպիրոխետներ:
Բակտերիաների կառուցվածքը
Իրենց կազմակերպվածության պարզությամբ և աննշան չափերով բակտերիաները պատկանում են ամենատարրական էակներին և կանգնած են կյանքի ամենացածր աստիճանների վրա: Չնայած գիտության և տեխնիկայի հսկայական առաջընթացին, բակտերիաների կառուցվածքի ոչ բոլոր հարցերը դեռ լուծված են:
Բակտերիաների մարմինը բաղկացած է էթիկական բովանդակության պատյանից և պրոտոպլազմայից՝ ներծծված բջջային հյութով։ Բակտերիաների թաղանթը բարակ է, անգույն, նրա կառուցվածքը մանրադիտակի տակ չի տարբերվում։ Դա տեսնելու համար դիմում են մշակման արհեստական մեթոդների։ Ծածկույթը ընկած է բջջի արտաքին ձևի հիմքում և, կարծես, հայտնի պաշտպանություն է անբարենպաստ պայմաններից: Ազատորեն պարուրելով բջջի պարունակությունը՝ շնորհիվ իր առաձգականության, այն թույլ է տալիս բակտերիաների ազատ տեղաշարժը, որը հաճախ ուղեկցվում է ամբողջ մարմնի արագ շարժումներով:
Կեղևի արտաքին շերտերը, ներծծելով ջուրը, հաճախ ուռչում են և կազմում դոնդողանման կպչուն զանգված՝ հասնելով նկատելի չափի։ Քանի որ արտաքին շերտերը դառնում են լորձաթաղանթ, կեղևը շարունակաբար նորանում է պրոտոպլազմայի հաշվին։ Սառեցված կպչուն պատյանը կոչվում է պարկուճ: Լորձի և պարկուճների ձևավորման ինտենսիվությունը կախված է սնուցման առանձնահատկություններից և երբեմն կարող է լինել շատ նշանակալի: Լորձաթաղանթային պարկուճը ձևավորվում է ոչ միայն յուրաքանչյուր բջջի մոտ առանձին-առանձին, այլ նաև բազմաթիվ բջիջներում, որոնք կապված են մեկ գաղութի մեջ և պարփակված, կարծես, ընդհանուր պարկուճում։ Բակտերիաների այս ցեխոտ գաղութները կոչվում են zoogles. Պարկուճների առաջացումը բնորոշ չէ բոլոր տեսակի բակտերիաներին։
բակտերիաների շարժում
Ինքնաբուխ շարժման ունակությունը բնորոշ է միայն բակտերիաների որոշ խմբերին: Բակտերիաները շարժվում են դրոշակի կամ թարթիչի միջոցով։ Դրոշակները նման են քիչ թե շատ երկար թելերի: Նրանք շատ նուրբ են, բարակ, հեշտությամբ կոտրվում են և տեսանելի չեն մանրադիտակի տակ առանց հատուկ ներկման: Նրանց տրամագիծը չի գերազանցում բակտերիալ մարմնի տրամագծի 1/20-ը։
Բակտերիաների շարժական ձևերը բաժանվում են հետևյալ խմբերի.
- միաձույլ - կա միայն մեկ բևեռային դրոշակ,
- lophotrichous - կա բջջի մի ծայրում դրոշակի կապոց,
- peritrichous flagella-ները գտնվում են մարմնի ողջ մակերեսով:
Դրոշակի տեղակայման բնույթը բակտերիայի մարմնի վրա որոշում է նրա շարժման բնույթը՝ ուղղագիծ կամ պատահական: Բակտերիաների շարժունակությունը կախված է մի շարք պայմաններից՝ ջերմաստիճանից, սննդային միջավայրի բաղադրությունից, նրանց կենսագործունեության արտադրանքներից և այլն։
սպորացում
Բազմաթիվ բակտերիաների մարմնում դրանց զարգացման որոշակի ժամանակահատվածներում հայտնվում են կլոր կամ էլիպսոիդ ձևավորումներ՝ հենարաններ։ Նրանք սովորաբար ավարտում են բակտերիաների զարգացման ցիկլը: Սպորների չափերը համեմատած բջիջների չափերի հետ, որոնք արտադրել են դրանք, կարող են շատ տարբեր լինել:
Բակտերիաների բոլոր տեսակների մոտ հենարաններ չեն ձևավորվում։ Նրանք շրջապատված են լավ մեկուսացված պատյանով, ջրի համար գրեթե անթափանց և ամենակայուն գոյացություններն են ողջ կենդանի աշխարհի մեջ: Այսպիսով, նրանք հաճախ դիմակայում են մի քանի ժամով եռման և չոր գոլորշու երկարատև ազդեցությանը 120-ից 140 ° ջերմաստիճանում: Շատ բացիլների սպորները պահպանում են իրենց կենսունակությունը երկար մնալուց հետո -190 ° ջերմաստիճանում և նույնիսկ հեղուկ ջրածնի ջերմաստիճանում (-253 °): Նրանք դիմացկուն են նաև քիմիական նյութերի` թույների ազդեցությանը: Այս ամենը չափազանց դժվարացնում է բակտերիաների պաթոգեն սպոր տեսակների դեմ պայքարը։
Հասուն սպորը կարող է կենսունակ մնալ տասնամյակներ շարունակ: Սովորաբար սպորների բողբոջումը տեղի է ունենում որոշակի քնած ժամանակահատվածից հետո և կապված է արտաքին պայմանների ազդեցության հետ։ Սպորացման ամբողջ գործընթացը տեղի է ունենում մեկ օրվա ընթացքում կամ ավելի քիչ: Սպորի հասունացումից հետո այն արտադրած բջիջը աստիճանաբար մահանում է, և հասուն սպորը դուրս է գալիս։ Ծլման ժամանակ ուռչում է, ջրով հարստանում, դրանից բարակ պատյան հագած սածիլ է դուրս գալիս։
Բակտերիաների վերարտադրություն
Հասնելով հասունության և աճի սահմանին, բակտերիաները սկսում են բազմանալ պարզ բաժանման միջոցով: Բաժանման ժամանակ մանրէի մարմնի միջին մասում առաջանում է միջնապատ, որն այնուհետ ճեղքվում է և բաժանում երկու նոր բջիջ։ Տարբեր բակտերիաների համար բաժանման ժամանակ միջնորմների հաջորդական դասավորությունը տարբեր է: Ձողաձև ձևերով այն գտնվում է երկար առանցքին ուղղահայաց, գնդաձև ձևերով միջնորմները կարող են տեղակայվել մեկ, երկու կամ երեք փոխադարձ ուղղահայաց հարթություններում, ինչը պատճառ է հանդիսանում այնպիսի ձևերի ձևավորման, ինչպիսիք են streptococci, tetracocci և sarcins: .
Բակտերիաների վերարտադրության արագությունը կախված է մի շարք պայմաններից և կարող է շատ տարբեր լինել: Որքան բարենպաստ են արտաքին պայմանները բակտերիաների գոյության համար, այնքան ավելի արագ է տեղի ունենում դրանց բաժանումը։ Նորմալ պայմաններում մանրէների թիվը կրկնապատկվում է մոտավորապես յուրաքանչյուր կես ժամը մեկ։ Եթե դա միշտ տեղի ունենար անխոչընդոտ, ապա մեկ բջջից բակտերիաների թիվը կհասներ հսկայական չափերի։ Մանրէաբան Կոնի կարծիքով, հինգ օրվա ընթացքում մեկ բացիլի սերունդը կարող է լցնել բոլոր ծովերն ու օվկիանոսները։ Այնուամենայնիվ, դա երբեք չի եղել և չի էլ լինելու: Բակտերիաների կյանքի ցիկլը սահմանափակվում է որոշակի արտաքին պայմաններով, որոնցից դուրս վերարտադրությունը դանդաղում է կամ ընդհանրապես դադարում: Սնուցման բացակայությունը, նյութափոխանակության վնասակար արտադրանքները, տարբեր տեսակների մրցակցությունը և այլն, վնասակար ազդեցություն են ունենում բակտերիաների վրա։ Անբարենպաստ պայմաններում նրանք զանգվածաբար մահանում են:
Բակտերիաների դասակարգում
Կենդանի էակների համակարգում բակտերիաների դիրքը դեռ լավ որոշված չէ։ Ընդհանրապես ընդունված է, որ բակտերիաները բույսերի աշխարհի մի մասն են, իսկ սնկերն ու ջրիմուռները նրանց ամենամոտ առնչվող օրգանիզմներն են։ Բակտերիաների մորֆոլոգիական առանձնահատկությունները շատ դեպքերում սահմանափակվում են մի քանի ձևերով՝ գնդաձև, ձողիկներ, պարույրներ: Նրանց արտաքին կազմակերպման արտասովոր պարզությունն ու տարրական բնույթը դժվարացնում են դրանք դասակարգելը։ Միայն մորֆոլոգիական բնութագրերի հիման վրա բակտերիաների տեսակը որոշելն անհնար է: Գիտական սիստեմատիկան հիմնված է մորֆոլոգիայի և զարգացման պատմության վրա, սակայն բակտերիաների դասակարգման համար անհրաժեշտ է իմանալ ոչ միայն մորֆոլոգիան, այլև դրանց ֆիզիոլոգիական և կենսաքիմիական բնութագրերը: Այս առումով սահմանվում են՝ բակտերիաների և թթվածնի հարաբերակցությունը, ջերմաստիճանի պայմանները, պիգմենտի առաջացումը, ժելատինի հեղուկացումը, շաքարների վրա թթուների և գազի առաջացումը, կաթի փոփոխությունը դրանում բակտերիաների աճի հետ, ինդոլի, ջրածնի սուլֆիդի, ամոնիակի առաջացումը, նիտրատների վերածումը նիտրիտների կամ ազատ ազոտի: Այնուամենայնիվ, սա միշտ չէ, որ բավարար է բակտերիաների տեսակը որոշելու համար:
Բակտերիաների դասակարգման տարբեր համակարգեր կան, բայց դրանք բոլորն էլ պայմանական են և շատ թե քիչ բնական դասակարգումից: Այս համակարգերի կամ գոնե դրանցից մեկի դիտարկումն այս դեպքում անհրաժեշտ չէ նույնիսկ ֆիտոպաթոգեն բակտերիաների նկատմամբ կիրառելիս: Պետք է միայն ասել, որ ներկայումս գրեթե բոլոր ֆիտոախտածին բակտերիաները միավորված են Pseudomonas, Xanthomonas, Bacterium եւ Erwinia սեռերում։
Վերջերս Մ. Վ. Գորլենկոն (1961) առաջարկեց Eubacteriales դասի ֆիտոպաթոգեն բակտերիաների դասակարգման հետևյալ համակարգը.
Ի. Mycobacteriaceae ընտանիքը(Չեսթեր, 1901) - ոչ շարժուն բակտերիաներ (առանց դրոշակի).
- 1-ին սեռ - Gorynebacterium (Leman and Neumann, 1896) - (գրամ դրական բակտերիաներ;
- 2-րդ սեռ Aplanobacterium (Smith, 1905, Geshich, 1956) - գրամ-բացասական բակտերիաներ:
II. Pseudomonadaceae ընտանիքը(Wilson et al., 1917) - բակտերիաներ flagella (բևեռային).
- 1-ին սեռ - Pseudomonas (Migula, 1900) - չբիծ և լյումինեսցենտ բակտերիաներ;
- 2-րդ սեռ - Xanthomonas (Dawson, 1839) - բակտերիաներ գունավոր գաղութներով:
III. Ընտանիք Bacteriaceae(Kon, 1872) - շարժական բակտերիաներ պերիտրիկ դրոշակներով, որոնք հենարաններ չեն կազմում.
- 1-ին սեռ - բակտերիա (Ehrenberger, 1828) - չներկված ձևեր, որոնք չեն ձևավորում պեկտինազներ և պրոտոպեկտինազներ.
- 2-րդ սեռ - Pectobacterium (Waldy, 1945) - չներկված ձևեր, որոնք ձևավորում են պեկտինազ և պրոտոպեկտինազ;
- 3-րդ սեռ - Chromobacterium (Bergonzini, 1881) - գունավոր ձևեր:
IV. Ընտանիք Bacillaceae(Ֆիշեր, 1895) - շարժական բակտերիաներ, սպոր ձևավորող ձողեր.
- 1-ին սեռ - Bacillus (Kon, 1832) - բջիջները չեն ուռչում կամ թեթևակի ուռչում սպորի ձևավորման ժամանակ.
- 2-րդ սեռ - Clostridium (Praznovsky, 1880) - բջիջները ուռչում են սպորի ձևավորման ժամանակ:
Վերոնշյալ համակարգում մինչ այժմ ընդհանուր ընդունված Erwinia ցեղը բաց է թողնված: Նրանից առանձնացված է հատուկ սեռ Pectobacterium, որը ներառում է բակտերիաներ՝ պերիտրիկ դրոշակակիր և պեկտոլիտիկ ակտիվությամբ։ Ֆիտոպաթոգեն բակտերիաներից նրանք, որոնք չունեն այս հատկությունը, վերագրվում են Bacterium սեռին: Այս համակարգը, ինքնին ռացիոնալ, նոր է և դեռ չի մտել առօրյա կյանք, հետևաբար, այս աշխատանքում մենք հավատարիմ ենք այն դասակարգմանը, որում իր տեղը տրված է Erwinia սեռին: Ֆիտոպաթոգեն բակտերիաների այս ընդհանուր անվանումը լայնորեն կիրառվում է մասնագիտացված գրականության մեջ ինչպես մեր երկրում, այնպես էլ արտերկրում։
Բակտերիաների տեսակը որոշելն առանց արհեստական սնուցող միջավայրի օգտագործման անհնար է: Այս առումով կարելի է նշել, որ մանրէներ մշակելիս նրանք կազմում են շատ բնորոշ գաղութներ։ Այս դեպքում բակտերիաների տեսակների մասին դատելու համար կարելի է օգտագործել մեկ տեսք։
Եթե սխալ եք գտնում, խնդրում ենք ընդգծել տեքստի մի հատվածը և սեղմել Ctrl+Enter.
1.5. Բակտերիաների ժամանակակից դասակարգումը Բակտերիաների ժամանակակից տաքսոնոմիայում ստեղծվել է մի իրավիճակ, որը բնորոշ է նաև այլ օրգանիզմների դասակարգմանը. տաքսոններ, սակայն արհեստական երևույթները պահպանում են իրենց նշանակությունը.– բնորոշ դասակարգումներ, ավելի հարմար միկրոօրգանիզմների նույնականացման համար։ Ներկայումս չկա պրոկարիոտների որևէ մանրամասն էվոլյուցիոն համակարգ և, ամենայն հավանականությամբ, այս խնդրի լուծումը մոտ ապագայի խնդիր չէ։ Մորֆոլոգիական, ֆիզիոլոգիական, կենսաքիմիական և գենետիկական կազմակերպման ոլորտում պրոկարիոտների առանձնահատկությունները ցույց են տալիս նրանց նկատմամբ բարձրակարգ օրգանիզմների համակարգի կառուցման մեջ օգտագործվող լավ զարգացած սկզբունքների անկիրառությունը: Չանդրադառնալով բակտերիաների տաքսոնոմիայի խնդրի պատմական ասպեկտներին, պետք է նշել, որ պրոկարիոտների համար առավել ընդունելի ֆիլոգենետիկ դասակարգման համակարգը համակարգն է, որը հիմնված է 16S rRNA-ում նուկլեոտիդային հաջորդականության համապատասխանության վրա: Այս համակարգը հիմք է հանդիսացել պրոկարիոտների բազմահատոր հանրագիտարանի 2-րդ հրատարակության համար. 26 ֆիլոգենետիկ «ճյուղեր» (խմբեր)՝ հիմնված իրենց 16S-rRNA կառուցվածքի վրա; 23 «ճյուղեր» ներկայացված են էվբակտերիայով, իսկ երեքը՝ արխեբակտերիայով։ Պետք է ընդգծել, որ այս ֆիլոգենետիկ խմբերի մեծ մասը պարունակում է պրոկարիոտ տեսակներ, որոնք չեն առանձնացվել որպես մաքուր կուլտուրաներ և, հետևաբար, դեռ մանրամասն ուսումնասիրված չեն: Այս տեսակների ներկայացուցիչների համար ներկայումս հայտնի են միայն 16S rRNA նուկլեոտիդային հաջորդականությունները։ Էվբակտերիաների 23 խմբերից երկու ֆիլոգենետիկ խմբերը ներկայացված են գրամ դրական բակտերիաներով, մնացած խմբերը գրամ-բացասական են։ Գրամ-բացասական բակտերիաները բաղկացած են պրոտեոբակտերիաների մեծ խմբից (Proteobacteria) և 20 խմբի այլ բակտերիաներից, որոնք ունեն այս տեսակի բջջային պատը: Պրոտեոբակտերիաների համառոտ նկարագրությունը, որոնց էուկարիոտների մեծ մասի միտոքոնդրիումները և քլորոպլաստները ամենամոտն են 16S-rRNA կազմով, տրված է Աղյուսակ 1-ում: 2. Պրոտեոբակտերիաները գրամ-բացասական բակտերիաների խումբ են, որոնք շատ տարասեռ են մորֆոլոգիական, ֆիզիոլոգիական և կենսաքիմիական առումներով: Այս խմբի ներկայացուցիչներին բնորոշ է բոլոր տեսակի էներգետիկ նյութափոխանակությունը և սնուցումը։ Պրոտեոբակտերիաների տեսակների մեծ մասի բջիջներն ունեն ձողաձև, գնդաձև կամ վիբրիոիդ ձև, դրանք բազմանում են հիմնականում երկուական տրոհման միջոցով, սակայն որոշ տեսակներ բնութագրվում են բողբոջումով և պտղաբեր մարմինների ձևավորմամբ բարդ բջջային ցիկլում: Այս խումբը պարունակում է ինչպես շարժական, այնպես էլ անշարժ բակտերիաներ, որոնք պայմանավորված են դրոշակներով: Ինչ վերաբերում է մոլեկուլային թթվածին, պրոտեոբակտերիաները դասակարգվում են որպես պարտադիր աերոբներ, պարտադիր և ֆակուլտատիվ անաէրոբներ: Proteobacteria խումբը բաժանված է հինգ ենթախմբի՝ հիմնված 16S rRNA-ի տարբերությունների վրա՝ ալֆա, բետա, գամմա, դելտա և էպսիլոն: Բացի պրոտեոբակտերիաներից, գրամ-բացասական էվբակտերիաները ներառում են էվբակտերիաների հետևյալ հիմնական խմբերը. և այլն: Գրամ-դրական բակտերիաների ֆիլոգենետիկ խմբեր - Actinobacteria և Firmicutes: Actinobacteria խումբը («ակտինոմիցետների ճյուղ») ներկայացված է ԴՆԹ-ում GC զույգերի բարձր պարունակությամբ բակտերիաների հետևյալ սեռերով. , Mycobacterium. Firmicutes խումբը («clostridial ճյուղ» - հիմնականում գրամ դրական բակտերիաներ, որոնք ունեն GC- զույգերի ցածր պարունակություն ԴՆԹ-ում) բաղկացած է հետևյալ սեռերից. , Sporosarcina, Bacillus, Desulfotomaculum, Heliobacterium, Mycoplasma, Ureaplasma եւ այլն: Archaebacteria-ն բաժանվում է երեք ֆիլոգենետիկ խմբի՝ Crenarchaeota, Euryarchaeota եւ Korarchaeota: Crenarchaeota խումբը բաղկացած է ծայրահեղ ջերմաֆիլ բակտերիաներից, որոնց ներկայացուցիչների մեծ մասը նյութափոխանակում է ծծումբը, ոմանք նվազեցնում են երկաթի և մոլիբդենի իոնները: Euryarchaeota խումբը ներառում է պարտադիր անաէրոբ մեթանոգեն արխեբակտերիաներ, ինչպես նաև ծայրահեղ ջերմաֆիլներ և հալոֆիլներ: Korarchaeota խումբը ձևավորվում է արխեբակտերիաներով, որոնք ապրում են ծծմբի տաք աղբյուրներում։ Մինչ օրս այս խմբի ներկայացուցիչներից և ոչ մեկը (ունենալով նմանատիպ 16S rRNA) մեկուսացված չէ որպես մաքուր մշակույթ, հետևաբար, նրանց ֆենոտիպիկ առանձնահատկությունները բավականաչափ ուսումնասիրված չեն: Ավարտելով պրոկարիոտների ֆիլոգենետիկ ճյուղերի դիտարկումը՝ պետք է նշել, որ առաջարկվող ֆիլոգենետիկ համակարգը, որը հիմնված է միայն մեկ ռիբոսոմային ՌՆԹ գենի նուկլեոտիդային հաջորդականությունների ուսումնասիրության վրա, ոչ այլ ինչ է, քան տեխնիկապես հարմար և զարգացած համակարգերից մեկը՝ բազմաթիվ պատվիրելու համար։ օրգանիզմներին՝ դրանք բացահայտելու համար, հետևաբար, տրամաբանական է կառուցել բակտերիաների ճիշտ դասակարգում, միայն հաշվի առնելով այս հատկանիշը հնարավոր չէ: Բակտերիաների ամենաճանաչված և օգտագործված ֆենոտիպային դասակարգումն այն է, որը ներկայացված է Burgey's Guide to Bacteria-ի իններորդ հրատարակությունում: Այս հրատարակության մեջ բակտերիաները բաժանվում են չորս հիմնական կատեգորիաների (բաժանումներ)՝ ելնելով բջջի սահմանային շերտի կառուցվածքից. 2) Firmicutes (լատ. firmus - ուժեղ) - գրամ դրական էվբակտերիաներ բջջային պատերով; 3) Tenericutes (լատիներեն tener - փափուկ, քնքուշ) - eubacteria, զուրկ բջջային պատերից; 4) Մենդոսիկուտներ (լատիներեն mendosus-ից՝ սխալ)՝ արխեբակտերիաներ, որոնց բջջային պատերը տարբերվում են այլ պրոկարիոտների նմանատիպ կառուցվածքներից։ Gracilicutes բաժանմունքը ներառում է տարբեր մորֆոլոգիայի բակտերիաներ՝ գրամ-բացասական բջջային պատով: Բազմացումը տեղի է ունենում հիմնականում երկուական տրոհման միջոցով, որոշ բակտերիաներ բազմանում են բողբոջելով։ Էնդոսպորը չի ձևավորվում: Շատերը շարժուն են. կան բակտերիաների շարժման բոլոր տեսակները՝ դրոշակների օգնությամբ, սահում, ծռում։ Բաժանումը ներառում է աերոբ, անաէրոբ և ֆակուլտատիվ անաէրոբ բակտերիաներ; ֆոտոտրոֆ և քիմոտրոֆ բակտերիաներ. Բաժանմունքը բաժանված է երեք դասի՝ Scotobacteria, Oxyphotobacteria, Anoxyphotobacteria: Scotobacteria դասը ներառում է գրամ-բացասական բակտերիաներ, որոնք չեն օգտագործում լույսի էներգիան նյութափոխանակության նպատակներով, այլ ստանում են միայն ռեդոքս ռեակցիաների արդյունքում։ Դասի անվանումը գալիս է հունարենից sсotos - խավար: Սա մանրէների ամենամեծ դասն է։ Անօքսիֆոտոբակտերիաների դասը ներառում է մանուշակագույն բակտերիաներ, կանաչ բակտերիաներ և հելիոբակտերիաներ, որոնք իրականացնում են անթթվածին ֆոտոսինթեզ (առանց մոլեկուլային թթվածնի արտազատման): Օքսիֆոտոբակտերիաների դասը ներկայացված է ցիանոբակտերիաներով և պրոքլորոֆիտներով, որոնք իրականացնում են թթվածնային ֆոտոսինթեզ (մոլեկուլային թթվածնի արտազատմամբ): Ֆոտոսինթեզի այս տեսակը նման է ֆոտոսինթեզի, որը տեղի է ունենում բույսերում։ Firmicutes բաժանմունքը ներառում է գրամ-դրական բջջային պատով բակտերիաներ: Բջիջները կարող են ունենալ տարբեր ձևեր՝ ձողիկներ, կոկիկներ, թելիկ, ճյուղավորվող։ Որոշ ներկայացուցիչներ ձևավորում են էնդոսպորներ։ Նրանցից շատերը անշարժ են. շարժուն ձևերն ունեն պերիտրիկ դրոշակներ: Բաժանմունքը ներառում է աերոբ, անաէրոբ և ֆակուլտատիվ անաէրոբ բակտերիաներ։ Բաժանմունքը բաղկացած է երկու դասից՝ Firmibacteria, Thallobacteria: Firmibacteria դասը ներառում է մեծ թվով «չճյուղավորված» գրամ դրական բակտերիաներ։ Thallobacteria դասը ներառում է բակտերիաներ, որոնց բջիջները ունակ են «ճյուղավորվել»: Tenericutes բաժանմունքը ներկայացված է բակտերիաներով, որոնք չունեն բջջային պատ։ Բջջային պատի բացակայության պատճառով բջիջների ձևը հաստատուն չէ. մեկ տեսակի մաքուր կուլտուրայում միաժամանակ առկա են կոկոիդ, ձողաձև, թելիկ, տանձաձև, սկավառակաձև և այլ բջիջներ: Այս բաժնում ընդգրկված բակտերիաների վերարտադրումը տեղի է ունենում երկուական տրոհման, բողբոջման միջոցով: Գրամ բիծը բացասական է: Բնորոշ է ագարի վերածվող փոքր գաղութների ձևավորումը։ Դրանք կարող են լինել սապրոֆիտ, մակաբույծ կամ ախտածին: Բաժանմունքը բաղկացած է մեկ դասի Mollicutes-ից (mycoplasmas): Mendosicutes բաժանումը ձևավորվում է կոշտ բջջային պատով բակտերիայով, բայց չունի մուրեին պեպտիդոգլիկան: Շատ ներկայացուցիչները խիստ անաէրոբներ են, որոնցից շատերը ունեն դրոշակ: Տեսակներին բնորոշ է էկոլոգիական և նյութափոխանակության բազմազանությունը, էքստրեմալ պայմաններում ապրելու կարողությունը։ Բաժանմունքը բաղկացած է մեկ դասից՝ Archaebacteria: Որպես չորս բաժանմունքների (հիմնական կատեգորիաների) մաս, հայտնաբերվել են բակտերիաների 35 խմբեր (կամ բաժիններ), որոնք այս կամ այն չափով կբնութագրվեն հաջորդ գլուխներում: Հետևյալ խմբերը պատկանում են Gracilicutes բաժնին. Խումբ 1. Սպիրոխետներ. Խումբ 2. Աերոբ (կամ միկրոաերոֆիլ), շարժուն, պարուրաձև (կամ վիբրիոիդ) Գրամ-բացասական բակտերիաներ։ Խումբ 3. Ոչ շարժուն կամ հազվադեպ շարժվող Գրամ-բացասական կոր բակտերիաներ: Խումբ 4. Գրամ-բացասական աերոբ (կամ միկրոաերոֆիլ) ձողեր և կոկկիներ: Խումբ 5. Ընտրովի aerobic գրամ-բացասական ձողեր. Խումբ 6. Գրամ-բացասական անաէրոբ ուղիղ, կոր կամ պարույր ձողեր: Խումբ 7. Ծծմբի կամ սուլֆատի դիսիմիլացիոն նվազեցում կատարող բակտերիաներ. Խումբ 8. Անաէրոբ գրամ-բացասական կոկիկներ. Խումբ 9. Ռիկեցիա և քլամիդիա. Խումբ 10. Անթթվածին ֆոտոտրոֆ բակտերիաներ. Խումբ 11. Թթվածնային ֆոտոտրոֆ բակտերիաներ. Խումբ 12. Աերոբ քիմոլիտոտրոֆ բակտերիաներ և հարակից օրգանիզմներ. Խումբ 13. Բողբոջող և (կամ) աճող բակտերիաներ. Խումբ 14. Բակտերիաներ պատյաններով. Խումբ 15. Ոչ ֆոտոսինթետիկ սահող բակտերիաներ, որոնք պտղաբեր մարմիններ չեն կազմում: Խումբ 16. Պտղատու մարմիններ կազմող սահող բակտերիաներ. Firmicutes բաժանմունքն ընդգրկում է. Խումբ 17. Գրամ դրական կոկիկներ. Խումբ 18. Գրամ-դրական ձողեր և կոկիներ, որոնք կազմում են էնդոսպորներ: Խումբ 19. Գրամ-դրական ճիշտ ձևի ձողեր, որոնք սպորներ չեն առաջացնում: Խումբ 20. Անկանոն ձևի գրամ-դրական ձողեր, որոնք սպորներ չեն առաջացնում: Խումբ 21. Միկոբակտերիա. 22–29 խմբեր. Ակտինոմիցետներ. Տեներիկուտների բաժանմունքին է պատկանում՝ 30-րդ խումբ. Միկոպլազմաներ. Division Mendosicutes-ը ներառում է՝ Խումբ 31. Մեթանոգեններ. Խումբ 32. Սուլֆատ վերականգնող արխեբակտերիաներ. Խումբ 33. Ծայրահեղ հալոֆիլ արխեբակտերիաներ (հալոբակտերիաներ). Խումբ 34. Բջջային պատից զուրկ արխեբակտերիաներ. Խումբ 35 Չափազանց ջերմասեր և հիպերթերմոֆիլ ծծմբի նյութափոխանակության արխեբակտերիաներ: Եզրափակելով, հարկ է ընդգծել, որ բնական համայնքներում գոյություն ունեցող միկրոօրգանիզմների մեծ մասը դեռևս կարիք ունի մեկուսացման մաքուր մշակույթների մեջ: Ենթադրվում է, որ ներկայումս ընդհանուր մանրէաբանական բազմազանության միայն 0,1%-ն է կարելի մշակել, և հնարավոր չէ աճեցնել և բացահայտել բակտերիաների մնացած ներկայացուցիչներին, թեև պրոկարիոտների մոտ 5 հազար տեսակ արդեն մեկուսացված և նկարագրված է մաքուր կուլտուրայում: