Klasifikacija mikroorganizama prema njihovim glavnim oblicima. Bakterije, njihova raznolikost. Struktura. Vitalnost. Osnovni pojmovi: bakterije
Ishrana bakterija.
Hrana.
pasivna difuzija
Olakšana difuzija
aktivni transport
U prvom slučaju, molekul nutrijenata formira kompleks sa proteinom periplazmatskog prostora koji je u interakciji sa specifičnom permeazom citoplazmatske membrane. Nakon energetski ovisnog prodiranja kroz citoplazmatsku membranu, kompleks "supstrat - periplazmatski protein - permeaza" disocira oslobađanjem molekula supstrata.
Tokom aktivnog transporta sa hemijskom modifikacijom transportovane supstance, lanac događaja uključuje: (1) fosforilaciju membranskog enzima-2 iz citoplazme pomoću fosfoenolpiruvata; (2) vezivanje na površini citoplazmatske membrane fosforilisanim molekulima supstrata enzima-2; (3) energetski ovisan transport molekula supstrata u citoplazmu; (4) transfer fosfatne grupe na molekul supstrata; (5) disocijacija kompleksa "supstrat-enzim" u citoplazmi. Zbog fosforilacije, molekuli supstrata se akumuliraju u citoplazmi stanica i ne mogu ih napustiti.
Klasifikacija bakterija prema vrsti hrane.
Putem unosa nutrijenata bakterije se svrstavaju u holofiti i Holozoic. Holofite bakterije (iz grčkog. holos- kompletan i phyticos- u vezi sa biljkama) nisu u stanju da ispuštaju u životnu sredinu enzime koji razgrađuju supstrate, usled čega hranljive materije troše isključivo u otopljenom, molekularnom obliku. Holozojske bakterije (iz grčkog. holos- kompletan i zoikos- u vezi sa životinjama), naprotiv, imaju kompleks egzoenzima koji obezbjeđuju vanjsku ishranu - razgradnju supstrata do molekula izvan bakterijske ćelije. Nakon toga, molekuli hranjivih tvari ulaze u unutrašnjost holozojske bakterije.
Po izvoru ugljika izolovani od bakterija autotrofi i heterotrofi. Autotrofi (iz grčkog. autos- sebe, trofej- hrana) ugljični dioksid (CO 2) se koristi kao izvor ugljika iz kojeg se sintetiziraju sve tvari koje sadrže ugljik. Za heterotrofi (od grčkog geteros - drugi i trofej- hrana) izvori ugljika su različite organske supstance u molekularnom obliku (ugljikohidrati, polihidrični alkoholi, aminokiseline, masne kiseline). Najveći stepen heterotrofije svojstven je prokariotima, koji mogu živjeti samo unutar drugih živih ćelija (na primjer, rikecije i klamidija).
Po izvoru dušika prokarioti se dijele u 3 grupe: 1) bakterije koje fiksiraju dušik (asimiliju molekularni dušik iz atmosferskog zraka); 2) bakterije koje troše neorganski azot iz amonijumovih soli, nitrita ili nitrata; 3) bakterije koje asimiliraju dušik sadržan u organskim jedinjenjima (aminokiseline, purini, pirimidini itd.).
Po izvoru energije bakterije se dijele na fototrofi i hemotrofi. fototrofne bakterije , kao i biljke, mogu koristiti sunčevu energiju. Fototrofni prokarioti ne uzrokuju bolesti kod ljudi. Hemotrofne bakterije primaju energiju u redoks reakcijama.
Po prirodi donora elektrona litotrofi(iz grčkog. litos- kamen) i organotrofi. At litotrofi (hemolitotrofi ) neorganske supstance djeluju kao donori elektrona (H 2, H 2 S, NH 3, sumpor, CO, Fe 2+ itd.). Donori elektrona organotrofi (hemoorganotrofi ) su organska jedinjenja - ugljeni hidrati, aminokiseline itd.
Većina bakterija patogenih za ljude ima hemoorganotrofni (hemoheterotrofni) tip ishrane; hemolitotrofni (hemoautotrofni) tip je rjeđi.
Sposobnošću sinteze organskih jedinjenja hemotrofne bakterije se dijele na prototrofi, auksotrofi i hipotrofi. prototrofne bakterije sintetiziraju sve potrebne organske tvari iz glukoze i amonijevih soli. Bakterije se zovu auksotrofi ako nisu u stanju sintetizirati bilo koju organsku supstancu iz navedenih spojeva. Ekstremni stepen gubitka metaboličke aktivnosti naziva se pothranjenost. hipotrofične bakterije obezbjeđuju svoju vitalnu aktivnost reorganizacijom struktura ili metabolita domaćina.
Osim ugljika i dušika, za punopravni život bakterija potrebni su ioni sumpora, fosfora i metala. Izvori sumpora su aminokiseline (cistein, metionin), vitamini, kofaktori (biotin, lipoična kiselina, itd.), sulfati. Izvori fosfora su nukleinske kiseline, fosfolipidi, fosfati. U dovoljno visokim koncentracijama, bakterijama je potreban magnezijum, kalij, kalcij, željezo; u znatno manjim količinama - cink, mangan, natrijum, molibden, bakar, nikl, kobalt.
faktori rasta- To su tvari koje bakterije ne mogu same sintetizirati, ali su im prijeko potrebne. Aminokiseline, azotne baze, vitamini, masne kiseline, gvožđe porfirini i druga jedinjenja mogu delovati kao faktori rasta. Da bi se stvorili optimalni uslovi za vitalnu aktivnost bakterija, hranljivim podlogama moraju se dodati faktori rasta.
Metabolizam, pretvaranje energije
A) Konstruktivni metabolizam.
Obavezna faza ishrane bakterija je asimilacija nutrijenata, odnosno njihovo uključivanje u izmijenjenom ili modificiranom obliku u sintetičke reakcije za reprodukciju staničnih komponenti i struktura.
Metabolizam proteina kod bakterija se može odvijati u 3 faze: primarni razgradnju proteina, sekundarni razgradnju proteina i sintezu proteina. Primarnu razgradnju proteinskih molekula do peptona vrše egzoenzimi - egzoproteaze koje bakterije oslobađaju u okoliš. Sekundarni propadanje nastaje pod dejstvom endoenzima (endoproteaza), koje imaju sve bakterije. Ovaj proces se odvija unutar bakterijske ćelije i sastoji se u razgradnji peptida na njihove sastavne aminokiseline. Potonji se mogu koristiti nepromijenjeni ili biti podvrgnuti hemijskim transformacijama (deaminacija, dekarboksilacija itd.), koje rezultiraju pojavom amonijaka, indola, sumporovodika, keto kiselina, alkohola, ugljičnog dioksida i dr. Detekcija ovih jedinjenja je od dijagnostičkog značaja u bakteriologiji.
Uz reakcije cijepanja proteina, javljaju se i reakcije njihove sinteze. Neke bakterije formiraju proteine iz gotovih aminokiselina dobivenih kao rezultat vanjske prehrane, dok druge bakterije samostalno sintetiziraju aminokiseline iz jednostavnih spojeva koji sadrže dušik i ugljik. Sinteza aminokiselina može se vršiti u reakcijama aminacije, transaminacije, amidacije, karboksilacije. Većina prokariota je u stanju sintetizirati sve aminokiseline koje čine ćelijske proteine. Karakteristika biosinteze aminokiselina je korištenje uobičajenih puteva biosinteze: ciklus trikarboksilne kiseline, glikoliza, oksidativni pentozofosfatni put, itd. Glavni početni spoj za sintezu aminokiselina su piruvat i fumarat.
metabolizam ugljikohidrata razlikuje se po autotrofima i heterotrofima (Shema 1). Autotrofne bakterije sintetiziraju sve potrebne ugljikohidrate iz ugljičnog dioksida. Sirovine za stvaranje ugljikohidrata u heterotrofnim bakterijama mogu biti: (1) jedno-, dvo- i trougljična jedinjenja; i (2) polisaharidi (skrob, glikogen, celuloza). Za razdvajanje potonjeg, mnoge heterotrofne bakterije imaju egzoenzime (amilaza, pektinaza, itd.), koji hidroliziraju polisaharide u glukozu, maltozu, fruktozu itd.
Kod autotrofnih bakterija, u Calvinovom ciklusu, iz ugljičnog dioksida nastaje ribuloza fosfat-fosfor-glicerinska kiselina, koja je uključena u reakcije glikolize koje idu u suprotnom smjeru. Konačni proizvod reverzne sinteze je glukoza.
Heterotrofne bakterije stvaraju glukozu iz jedinjenja s jednim, dva i tri ugljika, uključujući ih iu reakciju reverzne glikolize. Zbog činjenice da su neke reakcije glikolize ireverzibilne, heterotrofi su formirali posebne enzimske reakcije koje im omogućavaju da zaobiđu ireverzibilne reakcije kataboličkog puta.
Kada heterotrofne bakterije cijepaju polisaharide, nastali disaharidi ulaze u stanice i pod utjecajem maltoze, saharoze i laktoze podliježu hidrolizi i razgradnji na monosaharide, koji se zatim fermentiraju ili uključuju u reakcije međupretvorbe šećera.
metabolizam lipida. I egzogeni lipidi i amfiboliti intersticijalnog metabolizma mogu poslužiti kao polazni materijali za stvaranje lipida u bakterijama. Egzogeni lipidi su izloženi bakterijskim lipazama i drugim lipolitičkim enzimima. Mnoge vrste bakterija apsorbiraju glicerol, koji služi kao izvor plastičnog materijala i energije. Endogeni izvori za sintezu lipida mogu biti acetilkoenzim A, propionil-APB, malonil-APB (ACP - acetil-transportni protein), fosfodioksiaceton, itd.
Početni supstrat za sintezu masnih kiselina s parnim brojem atoma ugljika je acetilkoenzim A, za masne kiseline sa neparnim brojem atoma ugljika propionil-APB i malonil-APB. Stvaranje dvostrukih veza u molekulu kiseline u aerobnim prokariotima događa se uz sudjelovanje molekularnog kisika i enzima desaturaze. Kod anaerobnih prokariota, dvostruke veze se uvode u ranoj fazi sinteze kao rezultat reakcije dehidracije. Početni supstrat za sintezu fosfolipida je fosfodioksiaceton (međuspoj glikolitičkog puta), čija redukcija dovodi do stvaranja 3-fosforoglicerola. Zatim se ovoj drugoj dodaju 2 ostatka masnih kiselina u obliku kompleksa sa APB. Produkt reakcije je fosfatidna kiselina, čija aktivacija sa CTP-om i naknadnim vezivanjem na fosfatnu grupu serina, inozitola, glicerola ili nekog drugog spoja dovodi do sinteze odgovarajućih fosfolipida.
Mikroorganizmi koji su auksotrofni i hipotrofni za masne kiseline (na primjer, mikoplazme) dobivaju ih gotove iz stanica domaćina ili hranjivog medija.
Razmjena mononukleotida. Purinski i pirimidin mononukleotidi su bitne komponente DNK i RNK. Mnogi prokarioti su u stanju da koriste gotove purinske i pirimidinske baze, njihove nukleozide i nukleotide sadržane u hranljivom mediju, i da ih sintetiziraju iz supstanci male molekularne težine. Bakterije imaju enzime koji kataliziraju sljedeće faze interkonverzije egzogenih derivata purina i pirimidina: azotna baza - nukleozid - nukleotid (mono-di-trifosfat).
Sinteza purinskih i pirimidin mononukleotida de novo odvija na nezavisan način. Prilikom sinteze purinskih nukleotida, kao rezultat uzastopnih enzimskih reakcija, nastaje inozinska kiselina iz koje se kemijskim modifikacijama purinskog prstena sintetiziraju adenil (AMP) i guanilna (GMP) kiselina. Sinteza pirimidinskih nukleotida počinje stvaranjem orotidilne kiseline, čijom dekarboksilacijom nastaje uridilna kiselina (UMP). Od potonjeg nastaje UTP, čija acilacija dovodi do stvaranja CTP-a.
Deoksiribonukleotidi nastaju kao rezultat redukcije odgovarajućih ribonukleotida na nivou difosfata ili trifosfata. Sinteza nukleotida specifičnog za DNK, timidilne kiseline, odvija se enzimskom metilacijom deoksiuridilne kiseline.
Jonska izmjena. Mineralna jedinjenja - joni, NH 3 + , K + , Mg 2+ , Fe 2+ , SO 4 2- , PO 4 3- i druge bakterije dobijaju se iz okoline kako u slobodnom stanju tako iu stanju povezanom sa drugim organskim supstance. Kationi i anjoni se transportuju u bakterijsku ćeliju na različite načine, opisane u § 3. Na brzinu prodiranja jona u bakterijsku ćeliju utiče pH sredine i fiziološka aktivnost samih mikroorganizama.
B) Respiracija bakterija (energetski metabolizam).
Svi životni procesi su energetski zavisni, stoga je dobijanje energije izuzetno važan aspekt metabolizma prokariota. Energiju dobijaju anaerobnim i aerobnim disanjem.
Dah, ili biološka oksidacija je katabolički proces prijenosa elektrona sa donorske tvari na supstancu akceptor, praćen akumulacijom energije u makroergijskim jedinjenjima . Disanje se odvija u procesu kataboličkih reakcija, uslijed kojih složene organske tvari, cijepajući se, daju energiju i pretvaraju se u jednostavne spojeve. Energija akumulirana u makroergijskim supstancama (ATP, GTP, UTP, itd.) koristi se u anaboličkim reakcijama.
Prema načinu disanja mikroorganizmi se dijele na obavezni (strogi) aerobi, obvezni anaerobi i fakultativni anaerobi.
obavezni aerobi potreban slobodan kiseonik. Organska jedinjenja (ugljikohidrati, masti, proteini) su donori elektrona u hemoorganotrofnim aerobima patogenim za čoveka, a molekularni kiseonik je akceptor elektrona. Skladištenje energije u obliku ATP-a u hemoorganotrofnim aerobima događa se tokom oksidativne fosforilacije donora elektrona. Aerobi imaju citohrome (sudjeluju u prijenosu elektrona), kao i enzime (katalaza, superoksid dismutaza, peroksidaza) koji inaktiviraju toksične kisikove radikale koji nastaju tijekom disanja. Superoksid dismutaza inaktivira najtoksičniji metabolit, superoksidni radikal u H 2 O 2 . Enzim katalaza pretvara H 2 O 2 u H 2 O i O 2.
Posebna grupa aeroba su mikroaerofilne bakterije, koje, iako im je potreban kisik za energiju, bolje rastu s povećanim sadržajem CO 2, na primjer bakterije iz rodova Campylobacter i Helicobacter.
obavezni anaerobi ne trebaju slobodni kisik, naprotiv, čak i u malim količinama, kisik djeluje toksično na njih. Donori elektrona kod humanih patogenih anaerobnih hemoorganotrofa su različita organska jedinjenja (uglavnom ugljikohidrati). Akceptor elektrona u hemoorganotrofnim anaerobima su organska jedinjenja koja sadrže kiseonik - kiseline ili ketoni, odnosno akceptor elektrona je kiseonik povezan sa organskim fragmentom. Skladištenje energije u ovim prokariotima događa se tokom fosforilacije supstrata. Obavezni anaerobi, u pravilu, nemaju citokrome i enzime koji inaktiviraju kisikove radikale (negativni na katalazu i superoksid dismutazu).
U hemolitotrofnim anaerobima koji nisu patogeni za ljude, akceptor elektrona su anorganska jedinjenja koja sadrže kiseonik - nitrati, sulfati, karbonati.
Posebna grupa anaeroba su aerotolerantne bakterije koje mogu rasti u prisustvu atmosferskog kisika, ali ga ne koriste kao akceptor elektrona (na primjer, bakterije mliječne kiseline). Aerotolerantna katalaza i superoksid dismutaza prokarioti su pozitivni.
Fakultativni anaerobi mogu postojati i u kisikovim i u anoksičnim sredinama. Njihovi donori elektrona su organske supstance; Akceptori elektrona, u zavisnosti od uslova okoline, su molekularni ili kiseonik vezani u organskim i neorganskim jedinjenjima. Fakultativni anaerobi mogu akumulirati energiju i tokom oksidativne i supstratne fosforilacije. Poput aeroba, ova grupa bakterija ima citokrome i enzime antioksidativne odbrane.
Glavni supstrat za dobijanje energije su ugljikohidrati, koji se kod hemoheterotrofnih prokariota različitih tipova disanja mogu katabolizirati u acetil koenzim A („aktivirana octena kiselina“). Lipidi i proteini mogu djelovati kao energetski supstrati, budući da je acetilkoenzim A također jedan od međuproizvoda njihovog metabolizma (Shema 2).
Katabolizam ugljikohidrata kod hemoorganotrofnih prokariota uključuje: (a) anaerobne procese – glikolizu, pentozofosfatni put i ketodeoksifosfoglukonatni put; (b) aerobni proces - ciklus trikarboksilne kiseline (Krebsov ciklus). Anaerobni procesi se odvijaju kod svih prokariota, dok je aerobni proces karakterističan samo za obavezne aerobne i fakultativne anaerobne. Anaerobni putevi se zasnivaju na fosforilaciji supstrata, dok se aerobni procesi zasnivaju na oksidativnoj fosforilaciji.
Definicija pojmova.
Sterilizacija, dezinfekcija i antiseptici sastavni su dijelovi savremene medicinske, a posebno hirurške prakse. Razumijevanje principa i praktična primjena ovih metoda je od suštinskog značaja jer mnogi potencijalno patogeni mikroorganizmi mogu dugo preživjeti izvan domaćina, pokazati visoku otpornost na fizička i kemijska dezinficijensa i relativno lako se širiti s osobe na osobu.
Antiseptici- uništavanje ili sprečavanje rasta patogenih ili oportunističkih mikroorganizama hemijskim metodama. Ovaj izraz se obično koristi za vanjsku primjenu hemijskog preparata na živa tkiva.
antiseptik- supstanca koja inhibira rast ili uništava mikroorganizam (bez djelovanja na spore bakterija). Izraz je specifičan za supstance koje se koriste za lokalno djelovanje na živa tkiva.
Asepsa znači odsustvo sepse, ali općenito se ovaj izraz koristi da se naglasi odsustvo bilo kakvog živog organizma. Aseptičke metode označava svaki postupak koji je osmišljen da eliminira žive organizme i spriječi njihovu ponovnu kontaminaciju. Savremene hirurške i mikrobiološke tehnike zasnovane su na aseptičnim procedurama.
Biocid- supstanca koja ubija sve žive mikroorganizme, patogene i nepatogene, uključujući spore.
Biostat- sredstvo koje sprečava rast mikroorganizama, ali ih ne mora nužno i ubiti.
Dekontaminacija- uklanjanje mikroorganizama bez kvantitativnog određivanja. Ovaj termin je relativan; konačno uklanjanje mikroba može se postići sterilizacijom ili dezinfekcijom.
Dezinfekcija- proces koji smanjuje ili eliminira sve patogene osim spora.
Germicid- supstanca koja uništava mikroorganizme, posebno patogene. Germicid ne uništava spore.
Sanation- metoda kojom se mikrobna kontaminacija svodi na „siguran“ nivo. Ova metoda je ranije korištena za "pročišćavanje" neživih objekata.
Sterilizacija- upotreba fizičkih faktora i (ili) hemikalija za potpuno uništenje ili uništenje svih oblika mikrobnog života.
Sterilizacija.
Sterilizacija se definira kao uništavanje ili uklanjanje (filtracijom) svih mikroorganizama i njihovih spora. Sterilizacija se obično provodi toplinom. Sterilizacija, kao jedna od svakodnevnih rutina u radu mikrobiološke laboratorije, je suštinska metoda kojom se osigurava da kulture, oprema, pribor i podloga podržavaju rast samo potrebnih mikroorganizama, dok se ostali mikrobi uništavaju. Postoje takve vrste sterilizacije: kalcinacija u plamenu gorionika, prokuhavanje, djelovanje tekućom parom, para pod pritiskom u autoklavu, suha toplina, pasterizacija, tindalizacija, kemijska, hladna (mehanička) sterilizacija.
Izbor metoda sterilizacije.
Prilikom odabira metoda sterilizacije treba uzeti u obzir sljedeće zahtjeve:
1. Djelovanje: baktericidno, sporicidno, tuberkulocidno, fungicidno i virocidno.
2. Brzina postupka: sterilizaciju treba izvršiti što je brže moguće.
3. Permeabilnost: Sredstva za sterilizaciju moraju moći da prodru kroz pakovanje iu unutrašnjost instrumenta.
4. Kompatibilnost: ne bi trebalo biti promjena u strukturi ili funkciji materijala koji se steriliziraju nekoliko puta.
5. Netoksičnost: ne bi trebalo biti prijetnje ljudskom zdravlju i okolišu.
6. Postojanost organskog materijala: efikasnost sterilizacije ne bi trebalo da se smanji u prisustvu organskog materijala.
7. Prilagodljivost: mogućnost upotrebe za velike i male količine steriliziranog materijala.
8. Kontrola vremena: ciklus obrade mora se lako i precizno kontrolisati.
9. Cijena: razumna cijena opreme, instalacije i rada.
 Fizički sterilizatori
Fizički sterilizatori
mokro toplo, koji nastaje tokom procesa parnog autoklaviranja, glavno je sredstvo za sterilizaciju koje se koristi u kliničkim mikrobiološkim laboratorijama. Autoklavi se koriste za sterilizaciju medija kulture, vatrostalnih materijala i tretiranje infektivnog otpada. Parni sterilizator ili autoklav je izolirana komora pod pritiskom koja koristi zasićenu paru za stvaranje visokih temperatura (slika 1). Vazduh se uklanja iz komore pomeranjem mase ili vakuumom. Najčešće korišteni autoklavi sa zamjenom po težini. Lakša para se uvodi u komoru da istisne teži vazduh. Kratko izlaganje pari pod pritiskom može uništiti bakterijske spore. Za rutinsku sterilizaciju medija kulture i drugih materijala, vrijeme izlaganja je 15 minuta na 121°C i pritisku od 1,5 kg po 1 kvadratnom centimetru. Za infektivni otpad, vrijeme izlaganja se povećava na 30-60 minuta. Pored pravog vremena i temperature, za sterilizaciju je veoma bitan direktan kontakt sa parom. Prilikom rukovanja infektivnim materijalom treba osigurati maksimalan prodor pare u otpad. Takav materijal se mora obraditi na temperaturi od 132ºS. Antineoplastike, toksične hemikalije i radioizotope koji se ne mogu raspasti, kao i nestabilne hemikalije ne treba autoklavirati jer mogu ispariti i proširiti se kroz komoru kada su izloženi toploti.
 Sterilizacija suvom toplotom koristi se za materijale koji se ne mogu sterilizirati parom zbog mogućnosti oštećenja ili zbog nepropusnosti materijala za paru. Suva toplota je manje efikasna od vlažne i zahteva duže vreme izlaganja i više temperature. Sterilizacija suvom toplotom se obično izvodi u suvom toplotnom kabinetu (slika 2). Mehanizam sterilizacije suvom toplotom je oksidativni proces. Primeri materijala za koje se koristi sterilizacija suvom toplotom su ulja, praškovi, oštri instrumenti i stakleno posuđe. Suva toplota ili termička inaktivacija-sterilizacija koriste se kao alternativne metode za tretman infektivnog otpada.
Sterilizacija suvom toplotom koristi se za materijale koji se ne mogu sterilizirati parom zbog mogućnosti oštećenja ili zbog nepropusnosti materijala za paru. Suva toplota je manje efikasna od vlažne i zahteva duže vreme izlaganja i više temperature. Sterilizacija suvom toplotom se obično izvodi u suvom toplotnom kabinetu (slika 2). Mehanizam sterilizacije suvom toplotom je oksidativni proces. Primeri materijala za koje se koristi sterilizacija suvom toplotom su ulja, praškovi, oštri instrumenti i stakleno posuđe. Suva toplota ili termička inaktivacija-sterilizacija koriste se kao alternativne metode za tretman infektivnog otpada.
Pasterizacija uništava patogene brzim zagrijavanjem tvari do 71,1°C u trajanju od 15 sekundi, nakon čega slijedi brzo hlađenje. Pasterizacija nije sterilizacija jer nisu svi mikroorganizmi osjetljivi na nju. Ova metoda je eliminirala prijenos bolesti kao što su gastrointestinalna tuberkuloza i Q groznica putem hrane.
Tindalizacija je metoda intermitentne toplinske sterilizacije koja se može koristiti za ubijanje svih bakterija u otopinama. Budući da se rastuće bakterije lako ubijaju kratkim ključanjem (5 puta u roku od 1 sata po 5 minuta), sve što treba učiniti je ostaviti otopinu da odstoji određeno vrijeme prije nego što toplina poremeti sazrijevanje spora značajan gubitak njihove otpornosti na toplotu.
Filtracija je proces koji se koristi za uklanjanje mikroba i mikroskopskih čestica iz otopina, zraka i drugih plinova. Najčešća upotreba sterilizacije filtracijom u laboratoriji je za obradu dijagnostike, medija za kulturu, podloga za kulturu tkiva, seruma, otopina koje sadrže komponente seruma. Još jedna uobičajena primjena filtracije je sterilizacija zraka i plinova. Plastični ili papirni membranski filteri, koji se razlikuju po prečniku pora (od oko 12 do 0,22 µm) i koriste se za mehaničko odvajanje, služe i za prikupljanje mikroba iz tekućina za mikroskopsko ispitivanje ili kultivaciju direktno na filter kada se postavljaju na površinu natopljenu hranjivim tvarima. srednje.
ultraljubičasto zračenje je vrsta zračenja elektromagnetnih valova koja djeluje na ćelijsku nukleinsku kiselinu. Mikroorganizmi su vrlo osjetljivi na djelovanje ultraljubičastih zraka talasne dužine od 254 nm. Ultraljubičasto svjetlo se najčešće koristi za uništavanje mikroorganizama u zraku ili na površinama. Druge upotrebe su hladna sterilizacija određenih hemikalija i plastike za farmaceutske primene, sterilizacija seruma za ćelijske kulture i dezinfekcija vode. Značajan nedostatak ultraljubičastog zračenja kao sterilizatora je njegova nemogućnost prodiranja u materijale.
jonizujuće zračenje u elektromagnetnom spektru, ima smrtonosno dejstvo na mikroorganizme. Ovaj spektar uključuje mikrotalasne, γ-zrake, X-zrake i protok elektrona. Smrtonosni efekat jonizujućeg zračenja nastaje usled direktnog delovanja na ciljni molekul, usled čega se energija prenosi na molekul; a zbog indirektnog djelovanja - difuziju radikala.
ultrazvučna energija s niskom frekvencijom inaktivira mikroorganizme u vodenim otopinama. Fizički učinak sonikacije je posljedica kavitacije. Ultrazvučni čistači i drugi uređaji često se koriste za čišćenje instrumenata, ali se ne smatraju sterilizatorima. Međutim, kombinacija ultrazvuka i hemijskog tretmana ubija mikroorganizme.
Hemijski sterilizatori
2 % glutaraldehid Kao tečni hemijski sterilizator, ranije se široko koristio za obradu medicinskog i hirurškog materijala koji se ne može sterilisati grejanjem ili zračenjem. Glutaraldehid se takođe koristi u pripremi vakcina.
Dezinfekcija.
Dezinfekcija se može izvesti hemijskim metodama ili prokuhavanjem. Kuhanje je efikasna metoda za dezinfekciju instrumenata kao što su igle i špricevi ako autoklav nije dostupan. Prethodno očišćeni medicinski instrumenti treba da se kuvaju 20 minuta. Hemijska dezinfekcija se koristi za opremu osjetljivu na toplinu koja se može oštetiti visokim temperaturama. Hemijska sredstva za dezinfekciju kao što su komponente hlora, etil i izopropil alkohol, kvarterne amonijumske komponente i glutaraldehid se široko koriste.
Hemijska dezinfekciona sredstva.
Alkohol (etil i izopropil), rastvoren u vodi do koncentracije od 60-85%, veoma je efikasan u dezinfekciji. Alkoholi su baktericidni, fungicidni i tuberkulocidni, ali ne utiču na spore. Etil alkohol ima širi spektar virocidnog djelovanja od izopropil alkohola, pa je učinkovitiji protiv lipofilnih i hidrofilnih virusa.
Rješenje 37% formaldehid, koji se naziva formalin, može se koristiti kao sterilizator, dok se njegove koncentracije od 3-8% mogu koristiti kao dezinficijens.
fenol u svom čistom obliku ne koristi se kao dezinficijens zbog svoje toksičnosti, sposobnosti izazivanja razvoja tumora i korozije. Derivati fenola, u kojima funkcionalna grupa (hlor, brom, alkil, benzil, fenil, amil) zamjenjuje jedan od atoma vodika u aromatičnom prstenu, široko se koriste kao dezinficijensi. Ova zamjena smanjuje nedostatke fenola. Komponente fenola ubijaju mikrobe zbog inaktivacije enzimskih sistema, precipitacije proteina i narušavanja ćelijskog zida i membrane. Obično se koriste koncentracije od 2-5%, manje koncentracije zahtijevaju duže izlaganje.
Halogeni. Za dezinfekciju u laboratorijskoj praksi koriste se samo hlor i jod. Zbog činjenice da je hlor moćno oksidaciono sredstvo, vjeruje se da ubija mikrobe oksidacijom. Vjeruje se da jod ubija mikroorganizme reakcijom sa N-H i S-H grupama aminokiselina, kao i sa fenolnom grupom aminokiseline tirozin i dvostrukim vezama ugljik-ugljik nezasićenih masnih kiselina. Konvencionalni tretman uključuje prskanje 2-5% rastvora formaldehida u prisustvu pare na temperaturi od 60-80ºC.
Antiseptici.
Antiseptici se mogu naći u mikrobiološkim laboratorijama, prvenstveno u supstancama koje se koriste za pranje ruku. U slučajevima kada medicinsko osoblje pruža hitnu pomoć pacijentima koristeći supstance koje sadrže antibakterijske agense, to smanjuje broj bolničkih infekcija. Najčešće hemikalije koje se nalaze u pranju ruku su alkoholi, klorheksidin glukonat, jodofori, hloroksilenol i triklosan.
Tradicionalne metode tretiranja otpada i smeća su spaljivanje i parna sterilizacija.
Burning je metoda izbora za rukovanje otpadom i ostacima. Ova metoda čini otpad neinfektivnim i također mijenja njegov oblik i veličinu. Sterilizacija je efikasan način tretmana otpada, ali ne mijenja njegov oblik. Sterilizacija parom u autoklavu na 121°C u trajanju od najmanje 15 minuta ubija sve oblike mikrobnog života, uključujući veliki broj bakterijskih spora. Ova vrsta potpune sterilizacije može se obaviti i suvom toplotom na 160-170ºC u trajanju od 2-4 sata. Međutim, mora se osigurati da suva toplota bude u kontaktu sa materijalom koji se steriliše. Zbog toga se boce koje sadrže tečnost moraju labavo zatvoriti čepom ili pamučnim štapićima kako bi se para i toplota mogle razmenjivati sa vazduhom u bocama. Kontejneri za biološku opasnost koji sadrže otpad trebaju biti čvrsto vezani. Sterilizovani biološki opasni materijal mora biti zatvoren u odgovarajuće označene kontejnere.
Sterilizacija parom (autoklav). Zarazno smeće se smatra dekontaminiranim kada se broj vegetativnih bakterija, gljivica, mikobakterija i virusa koji sadrže lipide smanji za 6 lg puta, a bakterijskih endospora za 4 lg puta.
Ishrana bakterija.
Hrana. Pod ishranom bakterijske ćelije treba razumeti proces apsorpcije i asimilacije plastičnog materijala i energije kao rezultat transformativnih reakcija. . Vrste ishrane prokariota su složene i raznolike. Razlikuju se ovisno o načinu na koji hranjive tvari ulaze u bakterijsku ćeliju, izvorima ugljika i dušika, načinu dobivanja energije i prirodi donora elektrona.
transport hranljivih materija u ćeliju može se izvesti pomoću 3 mehanizma: pasivne difuzije, olakšane difuzije i aktivnog transporta.
pasivna difuzija je nespecifičan energetski ovisan proces koji se odvija duž koncentracijskog gradijenta supstanci (tvar iz sredine sa višom koncentracijom pasivno, prema zakonima osmoze, ulazi u sredinu sa nižom koncentracijom). Pasivnom difuzijom u bakterijsku ćeliju ulazi ograničena količina tvari, neki ioni, monosaharidi. Brzina prenosa supstanci tokom pasivne difuzije je neznatna i zavisi od lipofilnosti i veličine transportovanih molekula.
Olakšana difuzija je neisparljiv transport tvari duž gradijenta koncentracije uz pomoć enzima permeaze. Permeaze su specifični membranski proteini koji olakšavaju prolazak tvari kroz citoplazmatsku membranu. Permeaza fiksira na sebe molekul prenesene supstance, zajedno s kojom prevladava citoplazmatsku membranu, nakon čega se kompleks "tvar-permeaza" disocira. Oslobođena permeaza se koristi za provođenje drugih molekula. Kod prokariota, samo glicerol ulazi u ćeliju olakšanom difuzijom. U ovom slučaju, unutarćelijska koncentracija glicerola odgovara onoj izvan ćelije. Olakšana difuzija je najkarakterističnija za eukariotske mikroorganizme.
aktivni transport- ovo je energetski ovisan prijenos tvari u ćeliju protiv gradijenta koncentracije uz pomoć specifičnih enzima. Ogromna većina supstanci (joni, ugljikohidrati, aminokiseline, lipidi itd.) aktivnim transportom ulazi u bakterijsku ćeliju. Aktivni transport se može odvijati: (1) bez hemijske modifikacije transportovane supstance; (2) sa hemijskom modifikacijom.
Bakterije su prokariotski mikroorganizmi ćelijske strukture. Njihove veličine su od 0,1 do 30 mikrona. Mikrobi su izuzetno česti. Žive u tlu, vazduhu, vodi, snijegu, pa čak i toplim izvorima, na tijelu životinja, kao i unutar živih organizama, uključujući i ljudsko tijelo.
Raspodjela bakterija na vrste temelji se na nekoliko kriterija, među kojima se najčešće uzima u obzir oblik mikroorganizama i njihov prostorni raspored. Dakle, prema obliku ćelija, bakterije se dijele na:
Kokusi - mikro-, diplo-, strepto-, stafilokoki, kao i sarcini;
U obliku štapa - monobakterije, diplobakterije i streptobakterije;
Uvijene vrste - vibrioni i spirohete.
Bergeyeva determinanta sistematizira sve poznate bakterije prema principima identifikacije bakterija koje su našle najširu rasprostranjenost u praktičnoj bakteriologiji, na osnovu razlika u strukturi ćelijskog zida iu odnosu na Gramovu boju. Opis bakterija je dat po grupama (odjeljcima), koje uključuju porodice, rodove i vrste; u nekim slučajevima, grupe uključuju klase i redove. Bakterije patogene za ljude su uključene u mali broj grupa.
Ključ razlikuje četiri glavne kategorije bakterija -
Gracillicutes [od lat. gracilis, graciozan, tanak, + cutis, koža] - vrsta s tankim ćelijskim zidom, obojena gram negativan;
firmicutes [od lat. flrmus, jak, + cutis, koža] - bakterije sa debelim ćelijskim zidom, bojenje gram pozitivni;
Tenerikuti [od lat. tener, nježan, + cutis, koža] - bakterije koje nemaju ćelijski zid(mikoplazme i drugi pripadnici klase Mollicutes)
Mendosicutes [od lat. mendosus, nepravilan, + kutis, koža] - arhebakterije (reducirajuće metan i sulfat, halofilne, termofilne i arhebakterije, bez ćelijskog zida).
Grupa 2 Burgeyeva determinanta. Aerobne i mikroaerofilne pokretne uvijene i zakrivljene Gram-negativne bakterije. Vrste patogene za ljude su uključene u rodove Campylobacter, Helicobacters Spirillum.
Grupa 3 Bergeyeve determinante. Nepokretne (rijetko pokretne) Gram-negativne bakterije. Ne sadrži patogene vrste.
Grupa 4 Burgeyeve determinante. Gram-negativne aerobne i mikroaerofilne šipke i koke. Vrste patogene za ljude spadaju u porodice Legionellaceae, Neisseriaceae i Pseudomonada-ceae, u grupu spadaju i patogene i oportunističke bakterije iz rodova Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Moraxella, Kingella i
Grupa 5 Bergeyeve determinante. Fakultativni anaerobni gram-negativni štapići. Grupu čine tri porodice - Enterobacteriaceae, Vibrionaceae i Pasteurellaceae, od kojih svaka uključuje patogene vrste, kao i patogene i oportunističke bakterije iz rodova Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella i Streptobacillus.
Grupa 6 Bergeyeve determinante. Gram-negativne anaerobne ravne, zakrivljene i spiralne bakterije. Patogene i oportunističke vrste ubrajaju se u rodove Bacteroides, Fusobacterium, Porphoromonas i Prevotelta.
Grupa 7 Bergeyeve determinante. Bakterije koje vrše redukciju disimilacije sulfata ili sumpora Ne uključuje patogene vrste.
Grupa 8 Bergeyeve determinante. Anaerobne gram-negativne koke. Uključuje oportunističke bakterije iz roda Veillonella.
Grupa 9 Bergeyeve determinante. Rikecije i klamidija. Tri porodice - Rickettsiaceae, Bartonellaceae i Chlamydiaceae, od kojih svaka sadrži vrste patogene za ljude.
Grupe 10 i 11 Burgeyjevog vodiča uključuju anoksi- i kisikove fototrofne bakterije koje nisu patogene za ljude.
Grupa 12 Burgeyeve determinante. Aerobne hemolitotrofne bakterije i srodni organizmi. Kombinira bakterije koje oksidiraju i nitrifikuju sumpor, željezo i mangan, a koje ne oštećuju ljude.
Grupe 13 i 14 Burgeyjevog vodiča uključuju bakterije koje pupaju i/ili izrastaju i bakterije koje stvaraju ovojnicu. Predstavljaju ga vrste koje slobodno žive, nisu patogene za ljude;
Grupe 15 i 16 Burgeyjevog vodiča objedinjuju klizeće bakterije koje ne formiraju plodna tijela i formiraju ih. Grupe ne uključuju vrste patogene za ljude.
Grupa 17 Burgeyeve determinante. Gram-pozitivne koke. Uključuje oportunističke vrste iz rodova Enterococcus Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus.
Grupa 18 Burgeyeve determinante. Gram-pozitivni štapići i koke koje stvaraju spore. Uključuje patogene, uslovno patogene štapiće iz rodova Clostridium i Bacillus.
Grupa 19 Burgeyeve determinante. Gram-pozitivni štapići pravilnog oblika koji formiraju spore. Uključujući oportunističke vrste iz rodova Erysipelothrix i Listeria.
Grupa 20 Burgeyeve determinante. Gram-pozitivni štapići nepravilnog oblika koji formiraju spore. Grupa uključuje patogene i oportunističke vrste iz rodova Actinomyces, Corynebacterium Gardnerella, Mobiluncus itd.
Grupa 21 Burgeyeve determinante. Mycobacteria. Uključuje jedini rod Mycobacterium, koji kombinuje patogene i oportunističke vrste.
Grupe 22-29. Actinomycetes. Među brojnim vrstama, samo nokardioformni aktinomiceti (Grupa 22) iz rodova Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi i Terrabacter mogu izazvati lezije kod ljudi.
Grupa 30 Burgeyeve determinante. mikoplazme. Vrste uključene u rod Acholeplasma, Mycoplasma i Ureaplasma su patogene za ljude.
Preostale grupe Bergeyeve determinante - metanogene bakterije (31), sulfat-reducirajuće bakterije (32 ekstremno halofilne aerobne arhebakterije (33), arhebakterije lišene ćelijskog zida (34), ekstremni termofili i hipertermofili, koji ne metaboliziraju sumpor (35) sadrže vrste patogene za ljude.
Bakterije su najstarija grupa organizama koja trenutno postoji na Zemlji. Prve bakterije su se vjerovatno pojavile prije više od 3,5 milijardi godina i skoro milijardu godina bile su jedina živa bića na našoj planeti. Budući da su to bili prvi predstavnici divljih životinja, njihovo tijelo je imalo primitivnu strukturu.
S vremenom je njihova struktura postala složenija, ali se i danas bakterije smatraju najprimitivnijim jednoćelijskim organizmima. Zanimljivo je da neke bakterije još uvijek zadržavaju primitivne karakteristike svojih drevnih predaka. Ovo se opaža kod bakterija koje žive u vrelim sumpornim izvorima i anoksičnim muljem na dnu rezervoara.
Većina bakterija je bezbojna. Samo nekoliko je obojeno ljubičastom ili zelenom bojom. Ali kolonije mnogih bakterija imaju svijetlu boju, što je posljedica oslobađanja obojene tvari u okoliš ili pigmentacije stanica.
Otkrivač svijeta bakterija bio je Anthony Leeuwenhoek, holandski prirodnjak iz 17. stoljeća, koji je prvi stvorio savršeni mikroskop sa lupom koji povećava objekte 160-270 puta.
Bakterije su klasifikovane kao prokariote i odvojene su u posebno carstvo - Bakterije.
oblik tijela
Bakterije su brojni i raznoliki organizmi. Razlikuju se po formi.
| naziv bakterije | Oblik bakterija | Slika bakterije |
| cocci | sferni | |
| Bacillus |  | u obliku štapa |
| Vibrio | zakrivljeni zarez | |
| Spirillum |  | Spiralna |
| streptokoke |  | Lanac koka |
| Staphylococci |  | Grozdovi koka |
| diplococci | Dvije okrugle bakterije zatvorene u jednoj sluzavoj kapsuli |
Načini transporta
Među bakterijama postoje pokretni i nepokretni oblici. Pokretne se kreću talasastim kontrakcijama ili uz pomoć flagela (uvijenih spiralnih niti), koje se sastoje od posebnog proteina flagelina. Može biti jedna ili više flagela. Kod nekih bakterija se nalaze na jednom kraju ćelije, kod drugih - na dva ili po cijeloj površini.

Ali kretanje je svojstveno i mnogim drugim bakterijama koje nemaju flagele. Dakle, bakterije prekrivene sluzom izvana su sposobne kliziti.
Neke bakterije vode i tla bez flagela imaju plinske vakuole u citoplazmi. U ćeliji može biti 40-60 vakuola. Svaki od njih je ispunjen gasom (verovatno azotom). Regulacijom količine plina u vakuolama, vodene bakterije mogu potonuti u vodeni stupac ili izaći na njegovu površinu, dok se bakterije u tlu mogu kretati u kapilarama tla.
Stanište
Zbog jednostavnosti organizacije i nepretencioznosti, bakterije su široko rasprostranjene u prirodi. Bakterije se nalaze svuda: u kapi čak i najčistije izvorske vode, u zrncima zemlje, u vazduhu, na stenama, u polarnim snegovima, pustinjskom pesku, na dnu okeana, u ulju izvučenom iz velikih dubina, pa čak i u vrućim izvorska voda temperature oko 80ºS. Žive na biljkama, plodovima, kod raznih životinja i kod ljudi u crijevima, ustima, udovima i na površini tijela.
Bakterije su najmanja i najbrojnija živa bića. Zbog svoje male veličine, lako prodiru u sve pukotine, pukotine, pore. Veoma izdržljiv i prilagođen raznim uslovima postojanja. Podnose sušenje, ekstremnu hladnoću, zagrijavanje do 90ºS, bez gubitka vitalnosti.
Praktično ne postoji mjesto na Zemlji gdje se bakterije ne bi našle, ali u različitim količinama. Uslovi života bakterija su različiti. Nekima je potreban kiseonik iz vazduha, drugima ne treba i mogu da žive u okruženju bez kiseonika.
U zraku: bakterije se dižu u gornju atmosferu do 30 km. i više.
Posebno ih je mnogo u zemljištu. Jedan gram zemlje može sadržavati stotine miliona bakterija.
U vodi: u površinskim slojevima vode otvorenih rezervoara. Korisne vodene bakterije mineraliziraju organske ostatke.
U živim organizmima: patogene bakterije ulaze u organizam iz spoljašnje sredine, ali samo pod povoljnim uslovima izazivaju bolesti. Simbiotici žive u probavnim organima, pomažu u razgradnji i asimilaciji hrane, sintetiziraju vitamine.
Eksterna struktura
Bakterijska stanica je odjevena u posebnu gustu ljusku - staničnu stijenku, koja obavlja zaštitne i potporne funkcije, a također daje bakteriji trajni, karakterističan oblik. Ćelijski zid bakterije podseća na ljusku biljne ćelije. Propustljiv je: kroz njega hranjive tvari slobodno prolaze u ćeliju, a produkti metabolizma izlaze u okolinu. Bakterije često razvijaju dodatni zaštitni sloj sluzi, kapsulu, preko ćelijskog zida. Debljina kapsule može biti višestruko veća od prečnika same ćelije, ali može biti vrlo mala. Kapsula nije obavezan deo ćelije, ona se formira u zavisnosti od uslova u koje bakterija ulazi. Čuva bakterije od isušivanja.
Na površini nekih bakterija nalaze se duge flagele (jedna, dvije ili više) ili kratke tanke resice. Dužina flagele može biti višestruko veća od veličine tijela bakterije. Bakterije se kreću uz pomoć flagela i resica.

Unutrašnja struktura
Unutar bakterijske ćelije nalazi se gusta nepokretna citoplazma. Ima slojevitu strukturu, nema vakuola, pa se u samoj supstanci citoplazme nalaze razni proteini (enzimi) i rezervni nutrijenti. Bakterijske ćelije nemaju jezgro. U središnjem dijelu njihovih ćelija koncentrirana je supstanca koja nosi nasljedne informacije. Bakterije, - nukleinska kiselina - DNK. Ali ova supstanca nije uokvirena u jezgru.
Unutrašnja organizacija bakterijske ćelije je složena i ima svoje specifičnosti. Citoplazma je odvojena od ćelijskog zida citoplazmatskom membranom. U citoplazmi se razlikuju glavna tvar, ili matriks, ribosomi i mali broj membranskih struktura koje obavljaju različite funkcije (analozi mitohondrija, endoplazmatski retikulum, Golgijev aparat). Citoplazma bakterijskih stanica često sadrži granule različitih oblika i veličina. Granule mogu biti sastavljene od spojeva koji služe kao izvor energije i ugljika. Kapljice masti nalaze se iu bakterijskoj ćeliji.
U središnjem dijelu ćelije lokalizirana je nuklearna tvar, DNK, koja nije odvojena od citoplazme membranom. Ovo je analog jezgra - nukleoid. Nukleoid nema membranu, nukleolus i skup hromozoma.
Metode ishrane
Bakterije imaju različite načine hranjenja. Među njima su autotrofi i heterotrofi. Autotrofi su organizmi koji mogu samostalno formirati organske tvari za svoju ishranu.
Biljke trebaju dušik, ali same ne mogu apsorbirati dušik iz zraka. Neke bakterije kombinuju molekule dušika u zraku s drugim molekulima, što rezultira tvarima dostupnim biljkama.
Ove bakterije se naseljavaju u stanicama mladih korijena, što dovodi do stvaranja zadebljanja na korijenu, zvanih nodule. Takve kvržice nastaju na korijenu biljaka iz porodice mahunarki i nekih drugih biljaka.

Korijenje daje bakterijama ugljikohidrate, a bakterije korijenu daju tvari koje sadrže dušik koje biljka može preuzeti. Njihov odnos je obostrano koristan.
Korijeni biljaka luče mnoge organske tvari (šećere, aminokiseline i druge) kojima se hrane bakterije. Stoga se posebno mnoge bakterije naseljavaju u sloju tla koji okružuje korijenje. Ove bakterije pretvaraju mrtve biljne ostatke u supstance dostupne biljci. Ovaj sloj tla naziva se rizosfera.
Postoji nekoliko hipoteza o prodiranju bakterija kvržica u tkiva korijena:
- putem oštećenja epidermalnog i kortikalnog tkiva;
- kroz korijenske dlake;
- samo kroz mladu ćelijsku membranu;
- zbog pratećih bakterija koje proizvode pektinolitičke enzime;
- zbog stimulacije sinteze B-indoloctene kiseline iz triptofana, koja je uvijek prisutna u korijenskim izlučevinama biljaka.
Proces uvođenja nodulnih bakterija u tkivo korijena sastoji se od dvije faze:
- infekcija korijenskih dlačica;
- proces formiranja nodula.
U većini slučajeva, invazijska stanica se aktivno umnožava, formira takozvane infektivne niti i već u obliku takvih niti se kreće u biljna tkiva. Nodule bakterije koje su izašle iz niti infekcije nastavljaju da se razmnožavaju u tkivu domaćina.

Ispunjene brzomnožećim stanicama bakterija kvržica, biljne stanice počinju se intenzivno dijeliti. Veza mladog čvorića s korijenom mahunarke ostvaruje se zahvaljujući vaskularno-vlaknastim snopovima. U periodu funkcionisanja čvorovi su obično gusti. Do trenutka ispoljavanja optimalne aktivnosti, čvorići dobijaju ružičastu boju (zbog pigmenta legoglobina). Samo one bakterije koje sadrže legoglobin su sposobne da fiksiraju dušik.
Nodule bakterije stvaraju desetine i stotine kilograma dušičnih gnojiva po hektaru tla.
Metabolizam
Bakterije se međusobno razlikuju po metabolizmu. Za neke to ide uz učešće kiseonika, za druge - bez njegovog učešća.
Većina bakterija se hrani gotovim organskim tvarima. Samo nekoliko njih (plavo-zelene, ili cijanobakterije) u stanju je stvoriti organske tvari od neorganskih. Oni su igrali važnu ulogu u akumulaciji kiseonika u Zemljinoj atmosferi.
Bakterije upijaju tvari izvana, raskidaju njihove molekule, sastavljaju svoju ljusku iz ovih dijelova i nadopunjuju njihov sadržaj (tako rastu), a izbacuju nepotrebne molekule. Školjka i membrana bakterije omogućavaju joj da apsorbira samo prave tvari.

Kada bi ljuska i membrana bakterije bile potpuno nepropusne, nikakve tvari ne bi ušle u ćeliju. Kada bi bili propusni za sve supstance, sadržaj ćelije bi se pomešao sa medijumom – rastvorom u kojem bakterija živi. Za preživljavanje bakterija potrebna je ljuska koja propušta potrebne tvari, ali ne i one koje nisu potrebne.
Bakterija upija hranljive materije koje su joj blizu. Šta se dalje događa? Ako se može kretati samostalno (pomicanjem flageluma ili guranjem sluzi natrag), onda se kreće dok ne pronađe potrebne tvari.

Ako se ne može kretati, onda čeka dok mu difuzija (sposobnost molekula jedne tvari da prodre u gustinu molekula druge tvari) ne donese potrebne molekule.
Bakterije, zajedno sa drugim grupama mikroorganizama, obavljaju ogroman hemijski posao. Transformacijom različitih spojeva oni dobijaju energiju i hranjive tvari neophodne za njihovu vitalnu aktivnost. Metabolički procesi, načini dobivanja energije i potreba za materijalima za izgradnju tvari njihovog tijela u bakterijama su raznoliki.
Ostale bakterije zadovoljavaju sve potrebe za ugljikom neophodnim za sintezu organskih tvari tijela na račun neorganskih spojeva. Zovu se autotrofi. Autotrofne bakterije mogu sintetizirati organske tvari iz neorganskih. Među njima se ističu:
Hemosinteza
Upotreba energije zračenja je najvažniji, ali ne i jedini način stvaranja organske tvari iz ugljičnog dioksida i vode. Poznato je da bakterije ne koriste sunčevu svetlost kao izvor energije za takvu sintezu, već energiju hemijskih veza koje nastaju u ćelijama organizama tokom oksidacije određenih neorganskih jedinjenja - sumporovodika, sumpora, amonijaka, vodonika, azotne kiseline, jedinjenja gvožđa. gvožđa i mangana. Oni koriste organsku materiju koja se formira upotrebom ove hemijske energije za izgradnju ćelija svog tela. Stoga se ovaj proces naziva kemosinteza.
Najvažnija grupa hemosintetskih mikroorganizama su nitrificirajuće bakterije. Ove bakterije žive u tlu i vrše oksidaciju amonijaka, nastalog pri raspadanju organskih ostataka, do dušične kiseline. Potonji, reagirajući s mineralnim spojevima tla, pretvara se u soli dušične kiseline. Ovaj proces se odvija u dvije faze.
Bakterije željeza pretvaraju željezo željezo u oksid. Formirani željezni hidroksid se taloži i formira takozvanu močvarnu željeznu rudu.
Neki mikroorganizmi postoje zbog oksidacije molekularnog vodonika, čime se osigurava autotrofni način ishrane.
Karakteristična karakteristika vodikovih bakterija je sposobnost prelaska na heterotrofni način života kada su im snabdjeveni organski spojevi i u odsustvu vodika.
Dakle, kemoautotrofi su tipični autotrofi, jer samostalno sintetiziraju potrebna organska jedinjenja iz neorganskih supstanci, a ne uzimaju ih gotove od drugih organizama, poput heterotrofa. Hemoautotrofne bakterije razlikuju se od fototrofnih biljaka po potpunoj nezavisnosti od svjetlosti kao izvora energije.
bakterijska fotosinteza
Neke bakterije sumpora koje sadrže pigment (ljubičasta, zelena), koje sadrže specifične pigmente - bakteriohlorofile, sposobne su apsorbirati sunčevu energiju, uz pomoć koje se sumporovodik cijepa u njihovim organizmima i daje atome vodika za obnavljanje odgovarajućih spojeva. Ovaj proces ima mnogo zajedničkog s fotosintezom i razlikuje se samo po tome što je kod ljubičastih i zelenih bakterija sumporovodik (povremeno karboksilne kiseline) donor vodika, a kod zelenih biljaka to je voda. U tim i drugima, cijepanje i prijenos vodika se vrši zahvaljujući energiji apsorbiranih sunčevih zraka.
Takva bakterijska fotosinteza, koja se odvija bez oslobađanja kisika, naziva se fotoredukcija. Fotoredukcija ugljičnog dioksida povezana je s prijenosom vodika ne iz vode, već iz vodonik sulfida:
6CO 2 + 12H 2 S + hv → C6H 12 O 6 + 12S \u003d 6H 2 O
Biološki značaj kemosinteze i bakterijske fotosinteze na planetarnoj skali je relativno mali. Samo hemosintetske bakterije igraju značajnu ulogu u ciklusu sumpora u prirodi. Apsorbiran od strane zelenih biljaka u obliku soli sumporne kiseline, sumpor se obnavlja i postaje dio proteinskih molekula. Nadalje, tijekom uništavanja mrtvih biljnih i životinjskih ostataka od strane truležnih bakterija, sumpor se oslobađa u obliku sumporovodika, kojeg bakterije sumpora oksidiraju u slobodni sumpor (ili sumpornu kiselinu), koji stvara sulfite dostupne za biljke u tlu. Hemo- i fotoautotrofne bakterije su neophodne u ciklusu azota i sumpora.
sporulacija
Spore se formiraju unutar bakterijske ćelije. U procesu stvaranja spora, bakterijska ćelija prolazi kroz niz biohemijskih procesa. Količina slobodne vode u njoj se smanjuje, enzimska aktivnost se smanjuje. Time se osigurava otpornost spora na nepovoljne uvjete okoline (visoka temperatura, visoka koncentracija soli, sušenje, itd.). Formiranje spora je karakteristično za samo malu grupu bakterija.

Spore nisu bitna faza u životnom ciklusu bakterija. Sporulacija počinje tek nedostatkom nutrijenata ili nakupljanjem metaboličkih proizvoda. Bakterije u obliku spora mogu ostati u stanju mirovanja dugo vremena. Bakterijske spore podnose dugotrajno ključanje i veoma dugo zamrzavanje. Kada nastupe povoljni uslovi, spor klija i postaje održiv. Bakterijske spore su adaptacije za preživljavanje u nepovoljnim uslovima.
reprodukcija
Bakterije se razmnožavaju dijeljenjem jedne ćelije na dvije. Postigavši određenu veličinu, bakterija se dijeli na dvije identične bakterije. Tada se svaki od njih počinje hraniti, rasti, dijeliti i tako dalje.

Nakon produljenja ćelije postepeno se formira poprečni septum, a zatim ćelije kćeri divergiraju; kod mnogih bakterija, pod određenim uslovima, ćelije nakon deobe ostaju povezane u karakteristične grupe. U ovom slučaju, ovisno o smjeru ravnine podjele i broju podjela, nastaju različiti oblici. Razmnožavanje pupoljkom javlja se u bakterijama kao izuzetak.
Pod povoljnim uslovima, deoba ćelija kod mnogih bakterija se dešava svakih 20-30 minuta. Uz tako brzu reprodukciju, potomci jedne bakterije za 5 dana mogu formirati masu koja može ispuniti sva mora i okeane. Jednostavna računica pokazuje da se dnevno mogu formirati 72 generacije (720.000.000.000.000.000.000 ćelija). Ako se prevede u težinu - 4720 tona. Međutim, to se u prirodi ne događa, jer većina bakterija brzo umire pod utjecajem sunčeve svjetlosti, sušenja, nedostatka hrane, zagrijavanja na 65-100ºS, kao rezultat borbe između vrsta itd.
Bakterija (1), nakon što je apsorbirala dovoljno hrane, povećava se (2) i počinje se pripremati za reprodukciju (dioba ćelije). Njena DNK (u bakteriji je molekul DNK zatvoren u prsten) udvostručuje se (bakterija proizvodi kopiju ovog molekula). Čini se da su oba molekula DNK (3.4) pričvršćena za bakterijski zid i, kada se izduže, bakterije se razilaze sa strane (5.6). Prvo se dijeli nukleotid, a zatim citoplazma.
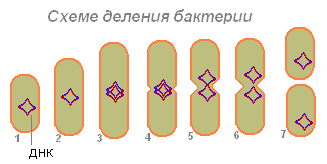
Nakon divergencije dva molekula DNK na bakterijama, pojavljuje se suženje, koje postepeno dijeli tijelo bakterije na dva dijela, od kojih svaki sadrži molekulu DNK (7).
Događa se (kod bacila sijena), dvije bakterije se slijepe i između njih se stvori most (1,2).

DNK se prenosi od jedne bakterije do druge preko skakača (3). Jednom u jednoj bakteriji, molekuli DNK se prepliću, na nekim mjestima se lijepe (4), nakon čega zamjenjuju dijelove (5).
Uloga bakterija u prirodi
Cirkulacija
Bakterije su najvažnija karika u općem kruženju tvari u prirodi. Biljke stvaraju složene organske tvari iz ugljičnog dioksida, vode i mineralnih soli tla. Ove tvari se vraćaju u tlo s mrtvim gljivama, biljkama i životinjskim leševima. Bakterije razlažu složene supstance na jednostavne, koje biljke ponovo koriste.
Bakterije uništavaju složenu organsku materiju mrtvih biljaka i životinjskih leševa, izlučevine živih organizama i razne otpadne materije. Hrane se ovim organskim supstancama, bakterije saprofitnog raspadanja pretvaraju ih u humus. To su oni redari naše planete. Dakle, bakterije su aktivno uključene u ciklus tvari u prirodi.
formiranje tla
Budući da su bakterije rasprostranjene gotovo posvuda i nalaze se u ogromnom broju, one u velikoj mjeri određuju različite procese koji se dešavaju u prirodi. U jesen lišće drveća i grmlja opada, nadzemni izdanci trave odumiru, stare grane otpadaju, a s vremena na vrijeme opadaju debla starih stabala. Sve se to postepeno pretvara u humus. U 1 cm 3. Površinski sloj šumskog tla sadrži stotine miliona saprofitnih bakterija tla nekoliko vrsta. Ove bakterije pretvaraju humus u različite minerale koje korijenje biljaka može apsorbirati iz tla.
Neke bakterije u tlu mogu apsorbirati dušik iz zraka, koristeći ga u životnim procesima. Ove bakterije koje fiksiraju dušik žive same ili se nastanjuju u korijenu mahunarki. Prodirući u korijenje mahunarki, ove bakterije uzrokuju rast korijenskih stanica i stvaranje kvržica na njima.
Ove bakterije oslobađaju dušikove spojeve koje biljke koriste. Bakterije dobijaju ugljene hidrate i mineralne soli iz biljaka. Dakle, postoji bliska veza između mahunarke i bakterija kvržica, što je korisno i za jedan i za drugi organizam. Ovaj fenomen se naziva simbioza.
Zahvaljujući simbiozi s bakterijama kvržica, mahunarke obogaćuju tlo dušikom, pomažući u povećanju prinosa.
Rasprostranjenost u prirodi
Mikroorganizmi su sveprisutni. Jedini izuzetak su krateri aktivnih vulkana i mala područja u epicentrima detoniranih atomskih bombi. Ni niske temperature Antarktika, ni kipući mlazovi gejzira, ni zasićene otopine soli u slanim bazenima, ni jaka insolacija planinskih vrhova, ni oštro zračenje nuklearnih reaktora ne ometaju postojanje i razvoj mikroflore. Sva živa bića su u stalnoj interakciji s mikroorganizmima, često ne samo njihova skladišta, već i distributeri. Mikroorganizmi su starosjedioci naše planete, aktivno razvijaju najnevjerovatnije prirodne supstrate.
Mikroflora tla
Broj bakterija u tlu je izuzetno velik - stotine miliona i milijarde jedinki u 1 gramu. Ima ih mnogo više u zemljištu nego u vodi i vazduhu. Ukupan broj bakterija u zemljištu varira. Broj bakterija ovisi o vrsti tla, njihovom stanju, dubini slojeva.
Na površini čestica tla mikroorganizmi se nalaze u malim mikrokolonijama (po 20-100 ćelija). Često se razvijaju u debljini ugrušaka organske tvari, na živim i umirućim korijenima biljaka, u tankim kapilarima i unutrašnjim grudvicama.
Mikroflora tla je veoma raznolika. Ovdje se nalaze različite fiziološke grupe bakterija: truležne, nitrifikujuće, azotno-fiksirajuće, sumporne bakterije itd. Među njima ima aerobnih i anaerobnih, spornih i nesporenih oblika. Mikroflora je jedan od faktora formiranja tla.
Područje razvoja mikroorganizama u tlu je zona uz korijenje živih biljaka. Zove se rizosfera, a ukupnost mikroorganizama sadržanih u njoj naziva se mikroflora rizosfere.
Mikroflora rezervoara
Voda je prirodno okruženje u kojem mikroorganizmi rastu u velikom broju. Većina ih ulazi u vodu iz tla. Faktor koji određuje broj bakterija u vodi, prisustvo nutrijenata u njoj. Najčistije su vode arteških bunara i izvora. Otvorene akumulacije i rijeke su veoma bogate bakterijama. Najveći broj bakterija nalazi se u površinskim slojevima vode, bliže obali. Sa povećanjem udaljenosti od obale i povećanjem dubine, broj bakterija se smanjuje.
Čista voda sadrži 100-200 bakterija po 1 ml, dok kontaminirana voda sadrži 100-300 hiljada ili više. Mnogo je bakterija u donjem mulju, posebno u površinskom sloju, gdje bakterije stvaraju film. U ovom filmu ima puno sumpornih i željeznih bakterija koje oksidiraju sumporovodik u sumpornu kiselinu i na taj način sprječavaju uginuće ribe. U mulju ima više sporonosnih oblika, dok u vodi preovlađuju oblici koji ne nose spore.
U pogledu sastava vrsta, vodena mikroflora je slična mikroflori tla, ali postoje i specifični oblici. Uništavajući razni otpad koji je pao u vodu, mikroorganizmi postepeno provode takozvano biološko pročišćavanje vode.
Mikroflora vazduha
Mikroflora vazduha je manje brojna od mikroflore zemlje i vode. Bakterije se uzdižu u zrak s prašinom, mogu tamo ostati neko vrijeme, a zatim se talože na površini zemlje i umiru od nedostatka ishrane ili pod uticajem ultraljubičastih zraka. Broj mikroorganizama u zraku ovisi o geografskom području, lokaciji, godišnjem dobu, zagađenosti prašinom itd. Svaka zrnca prašine je nosilac mikroorganizama. Većina bakterija u vazduhu iznad industrijskih preduzeća. Vazduh na selu je čistiji. Najčistiji vazduh je iznad šuma, planina, snežnih prostora. Gornji slojevi vazduha sadrže manje klica. U mikroflori zraka nalazi se mnogo pigmentiranih bakterija koje nose spore koje su otpornije od drugih na ultraljubičaste zrake.
Mikroflora ljudskog organizma
Tijelo čovjeka, čak i potpuno zdravog, uvijek je nosilac mikroflore. Kada ljudsko tijelo dođe u kontakt sa zrakom i tlom, različiti mikroorganizmi, uključujući patogene (bacile tetanusa, plinsku gangrena, itd.), talože se na odjeću i kožu. Izloženi dijelovi ljudskog tijela su najčešće kontaminirani. E. coli, stafilokoki se nalaze na rukama. U usnoj šupljini postoji preko 100 vrsta mikroba. Usta su svojom temperaturom, vlažnošću, ostacima hranljivih materija odlično okruženje za razvoj mikroorganizama.
Želudac ima kiselu reakciju, pa većina mikroorganizama u njemu umire. Počevši od tankog crijeva, reakcija postaje alkalna, tj. povoljno za mikrobe. Mikroflora u debelom crijevu je vrlo raznolika. Svaka odrasla osoba dnevno izluči oko 18 milijardi bakterija sa izmetom, tj. više pojedinaca nego ljudi na planeti.
Unutrašnji organi koji nisu povezani sa spoljašnjom sredinom (mozak, srce, jetra, bešika, itd.) obično su bez mikroba. Mikrobi ulaze u ove organe samo tokom bolesti.
Bakterije u biciklizmu
Mikroorganizmi općenito, a bakterije posebno igraju važnu ulogu u biološki važnim ciklusima materije na Zemlji, vršeći kemijske transformacije koje su potpuno nedostupne ni biljkama ni životinjama. Različite faze ciklusa elemenata provode organizmi različitih tipova. Postojanje svake posebne grupe organizama zavisi od hemijske transformacije elemenata koju vrše druge grupe.
ciklus azota
Ciklična transformacija azotnih jedinjenja igra glavnu ulogu u snabdevanju potrebnim oblicima azota različitim organizmima biosfere u smislu nutritivnih potreba. Preko 90% ukupne fiksacije dušika je zbog metaboličke aktivnosti određenih bakterija.
Ciklus ugljenika
Biološka transformacija organskog ugljika u ugljični dioksid, praćena redukcijom molekularnog kisika, zahtijeva zajedničku metaboličku aktivnost različitih mikroorganizama. Mnoge aerobne bakterije provode potpunu oksidaciju organskih tvari. U aerobnim uvjetima, organska jedinjenja se u početku razgrađuju fermentacijom, a organski krajnji proizvodi fermentacije se dalje oksidiraju anaerobnim disanjem ako su prisutni neorganski akceptori vodonika (nitrat, sulfat ili CO2).
Ciklus sumpora
Za žive organizme, sumpor je dostupan uglavnom u obliku rastvorljivih sulfata ili redukovanih organskih jedinjenja sumpora.
Ciklus gvožđa
Neki rezervoari slatke vode sadrže visoke koncentracije redukovanih soli željeza. Na takvim mjestima razvija se specifična bakterijska mikroflora – željezne bakterije, koje oksidiraju redukovano željezo. Učestvuju u stvaranju močvarnih željeznih ruda i izvora vode bogatih solima željeza.
Bakterije su najstariji organizmi, pojavili su se prije oko 3,5 milijardi godina u Arheju. Oko 2,5 milijardi godina dominirali su Zemljom, formirajući biosferu, i učestvovali u formiranju atmosfere kiseonika.
Bakterije su jedni od najjednostavnije uređenih živih organizama (osim virusa). Vjeruje se da su oni prvi organizmi koji su se pojavili na Zemlji.
bakterije- To su jednoćelijski organizmi bez hlorofila koji se vegetativno razmnožavaju diobom, rjeđe vezivanjem, ponekad formirajući unutarćelijske spore.
Veličina bakterija se mjeri u mikronima i, uz rijetke izuzetke, kreće se od 0,06-0,3 do 3-5 μ. Kap vode može lako da primi nekoliko stotina miliona bakterija.
Oblik bakterijske ćelije je prilično ujednačen. Poznata su tri glavna oblika bakterija: okrugli, štapićasti i uvijeni s brojnim i neupadljivim prijelazima između njih. Anton De Bari ih je figurativno uporedio s loptom za bilijar, olovkom i vadičepom.
Bakterije okruglog oblika nazivaju se kokije. Razlikuju se po veličini i relativnom položaju. Koke povezane u parove nazivaju se diplokoki, ali povezani u obliku ogrlice nazivaju se streptokoki. Prilikom dijeljenja, naizmjenično u dva međusobno okomita smjera, nastaju tetrakoki. Ako je podjela ispravna i ponavlja se u tri međusobno okomita smjera, tada se ćelijske veze formiraju u obliku paketa - to su takozvane sardine. Dijeleći se u različitim smjerovima bez puno pravilnosti, kokice formiraju nasumične grozdove nalik na četku grožđa. Zovu se stafilokoki.
Bakterije u obliku štapa nešto su raznolikije po svom izgledu. Mogu biti sa skraćenim ili zaobljenim krajevima, cilindrični, bačvasti ili limunasti i, takoreći, sa suženjem u sredini, često elipsoidni, koji se razlikuju samo po širini i dužini. Štapovi mogu biti ravni, zakrivljeni, pojedinačni, povezani u parove ili u lanac, kratki ili jako izduženi. Bakterije u obliku štapa, kod kojih je dužina dvostruko ili veća od širine, nazivaju se bacili; ako je razlika između dužine i širine mala, onda se nazivaju bakterijama.
Bakterije kovrčavog oblika razlikuju se ne samo po dužini i debljini, već i po broju i prirodi kovrča. Blago zakrivljene bakterije (kovrča ne prelazi 1/4 okreta spirale) nazivaju se vibrio, bakterije s jednom ili više velikih pravilnih kovrča nazivaju se spirila. Duge i tanke bakterije uvijenog oblika s brojnim sitnim uvojcima, ponekad s velikim zakrivljenjima cijelog filamenta, nazivaju se spirohete.
Struktura bakterija
Po jednostavnosti organizacije i neznatnoj veličini, bakterije spadaju u najelementarnija bića i stoje na najnižim stepenicama života. Uprkos ogromnom napretku nauke i tehnologije, sva pitanja o strukturi bakterija još uvek nisu rešena.
Tijelo bakterije sastoji se od ljuske i protoplazme etičkog sadržaja, impregniranog ćelijskim sokom. Ljuska bakterije je tanka, bezbojna, njena struktura se ne može razlikovati pod mikroskopom. Da bi to vidjeli, pribjegavaju umjetnim metodama obrade. Omotač je u osnovi spoljašnjeg oblika ćelije i čini se da je dobro poznata odbrana od nepovoljnih uslova. Slobodno obavija sadržaj ćelije, zahvaljujući svojoj elastičnosti, omogućava slobodno kretanje bakterija, često praćeno žustrim pokretima celog tela.
Vanjski slojevi ljuske, upijajući vodu, često nabubre i formiraju želatinoznu ljepljivu masu, dostižući primjetnu veličinu. Kako vanjski slojevi postaju sluzavi, ljuska se kontinuirano obnavlja na račun protoplazme. Ohlađena ljepljiva ljuska naziva se kapsula. Intenzitet stvaranja sluzi i kapsula zavisi od karakteristika ishrane i ponekad može biti veoma značajan. Sluzna kapsula se formira ne samo u blizini svake ćelije posebno, već iu mnogim ćelijama koje su povezane u jednu koloniju i zatvorene, takoreći, u zajedničku kapsulu. Ove sluzave kolonije bakterija nazivaju se zoogles. Formiranje kapsula nije karakteristično za sve vrste bakterija.
kretanje bakterija
Sposobnost spontanog kretanja svojstvena je samo nekim grupama bakterija. Bakterije se kreću pomoću flagela ili cilija. Flagele izgledaju kao manje ili više dugi filamenti. Vrlo su osjetljive, tanke, lako se lome i nisu vidljive pod mikroskopom bez posebnog bojenja. Njihov promjer ne prelazi 1/20 promjera tijela bakterije.
Mobilni oblici bakterija dijele se u sljedeće grupe:
- monotrih - postoji samo jedan polarni flagelum,
- lophotrichous - nalazi se snop flagela na jednom kraju ćelije,
- peritrihozne flagele nalaze se po cijeloj površini tijela.
Priroda položaja flagele na tijelu bakterije određuje prirodu njenog kretanja - pravolinijsko ili nasumično. Mobilnost bakterija zavisi od niza uslova: temperature, sastava hranljive podloge, proizvoda njihove vitalne aktivnosti itd. Pokretni oblici bakterija nisu opremljeni flagelama u svim fazama svog razvoja i ne u svim uslovima rasta.
sporulacija
U tijelu mnogih bakterija, u određenim periodima njihovog razvoja, pojavljuju se okrugle ili elipsoidne formacije - oslonci. Obično završavaju razvojni ciklus bakterija. Veličina spora u poređenju sa veličinom ćelija koje su ih proizvele može uveliko varirati.
Oslonci se ne formiraju kod svih vrsta bakterija. Okruženi su dobro izoliranom školjkom, gotovo nepropusnom za vodu i najstabilnije su formacije među cjelokupnim živim svijetom. Dakle, često izdržavaju nekoliko sati ključanja i produženo izlaganje suhoj pari na temperaturama od 120 do 140 °. Spore mnogih bacila zadržavaju svoju održivost nakon dugog boravka na temperaturi od -190 °, pa čak i na temperaturi tekućeg vodika (-253 °). Otporne su i na djelovanje hemikalija – otrova. Sve ovo izuzetno otežava borbu protiv patogenih spornih vrsta bakterija.
Zrela spora može ostati održiva decenijama. Obično se klijanje spora javlja nakon određenog perioda mirovanja i povezano je sa uticajem spoljašnjih uslova. Cijeli proces sporulacije odvija se u roku od jednog dana ili manje. Nakon što spora sazri, ćelija koja ju je proizvela postepeno odumire i zrela spora izlazi. Prilikom klijanja nabubri, postaje bogatiji vodom i iz njega izlazi sadnica obučena u tanku ljusku.
Reprodukcija bakterija
Nakon dostizanja granice zrelosti i rasta, bakterije se počinju razmnožavati jednostavnom dijeljenjem. Prilikom diobe u srednjem dijelu tijela bakterije pojavljuje se septum, koji se zatim razdvaja i razdvaja dvije nove ćelije. Sekvencijski raspored pregrada tokom podjele različit je za različite bakterije. Kod štapićastih oblika nalazi se okomito na dugu os, u sfernim oblicima pregrade se mogu nalaziti u jednoj, dvije ili tri međusobno okomite ravni, što je razlog za stvaranje oblika kao što su streptokoki, tetrakoki i sarcini. .
Brzina razmnožavanja bakterija ovisi o brojnim uvjetima i može biti vrlo različita. Što su vanjski uvjeti za postojanje bakterija povoljniji, to brže dolazi do njihove podjele. U normalnim uslovima, broj bakterija se udvostručuje otprilike svakih pola sata. Kada bi se to uvijek odvijalo nesmetano, tada bi broj bakterija iz jedne ćelije dostigao kolosalne razmjere. Prema mikrobiologu Kohnu, potomci jednog bacila za pet dana mogli bi ispuniti sva mora i okeane. Međutim, to se nikada nije dogodilo i nikada neće. Životni ciklus bakterija ograničen je određenim vanjskim uvjetima, nakon kojih se reprodukcija usporava ili potpuno zaustavlja. Nedostatak ishrane, štetni produkti metabolizma, konkurencija raznih vrsta itd., štetno utiču na bakterije. U nepovoljnim uslovima masovno umiru.
Klasifikacija bakterija
Položaj bakterija u sistemu živih bića još nije dobro definisan. Općenito je prihvaćeno da su bakterije dio biljnog svijeta, a gljive i alge su im najbliži organizmi. Morfološke karakteristike bakterija u većini slučajeva ograničene su na nekoliko oblika: sferni, štapićasti, spiralni. Izuzetna jednostavnost i elementarna priroda njihove vanjske organizacije otežavaju njihovu klasifikaciju. Odrediti vrstu bakterije samo na osnovu morfoloških karakteristika je nemoguće. Naučna sistematika se zasniva na morfologiji i istoriji razvoja, ali za klasifikaciju bakterija potrebno je poznavati ne samo morfologiju, već i njihove fiziološke i biohemijske karakteristike. S tim u vezi utvrđuju se: odnos bakterija i kiseonika, temperaturni uslovi, stvaranje pigmenta, ukapljivanje želatina, stvaranje kiselina i gasova na šećerima, promena mleka sa rastom bakterija u njemu, stvaranje indola, sumporovodika, amonijaka, redukcija nitrata u nitrite ili slobodni dušik. Međutim, to nije uvijek dovoljno za određivanje vrste bakterije.
Postoje različiti klasifikacijski sistemi za bakterije, ali su svi uslovni i manje-više od prirodne klasifikacije. Razmatranje ovih sistema ili barem jednog od njih u ovom slučaju nije potrebno čak ni kada se primjenjuju na fitopatogene bakterije. Treba samo reći da su trenutno gotovo sve fitopatogene bakterije objedinjene u rodove Pseudomonas, Xanthomonas, Bacterium i Erwinia.
Nedavno je M. V. Gorlenko (1961) predložio sljedeći sistem klasifikacije za fitopatogene bakterije klase Eubacteriales:
I. Porodica Mycobacteriaceae(Chester, 1901) - nepomične bakterije (bez flagela):
- 1. rod - Gorynebacterium (Leman i Neumann, 1896) - (gram-pozitivne bakterije;
- 2. rod Aplanobacterium (Smith, 1905, Geshich, 1956) - gram-negativne bakterije.
II. Porodica Pseudomonadaceae(Wilson et al., 1917) - bakterije sa flagelama (polarne):
- 1. rod - Pseudomonas (Migula, 1900) - neobojene i fluorescentne bakterije;
- 2. rod - Xanthomonas (Dawson, 1839) - bakterije sa obojenim kolonijama.
III. Porodica Bacteriaceae(Kon, 1872) - pokretne bakterije s peritrihoznim flagelama koje ne formiraju nosače:
- 1. rod - Bakterija (Ehrenberger, 1828) - neobojeni oblici koji ne stvaraju pektinaze i protopektinaze;
- 2. rod - Pectobacterium (Waldy, 1945) - neobojeni oblici koji formiraju pektinazu i protopektinazu;
- 3. rod - Chromobacterium (Bergonzini, 1881) - obojeni oblici.
IV. Porodica Bacillaceae(Fischer, 1895) - pokretne bakterije, štapići koji stvaraju spore:
- 1. rod - Bacillus (Kon, 1832) - ćelije ne bubre ili blago bubre tokom formiranja spora;
- 2. rod - Clostridium (Praznovsky, 1880) - ćelije bubre tokom formiranja spora.
U navedenom sistemu izostavljen je do sada opšteprihvaćeni rod Erwinia. Iz nje je izoliran poseban rod Pectobacterium, koji uključuje bakterije s peritrihoznim flagelama i pektolitičkim djelovanjem. One od fitopatogenih bakterija koje ne posjeduju ovu sposobnost pripisuju se rodu Bacterium. Ovaj sistem, racionalan sam po sebi, nov je i još nije ušao u svakodnevni život, stoga se u ovom radu pridržavamo klasifikacije u kojoj je rod Erwinia dobio svoje mjesto. Ovaj generički naziv fitopatogenih bakterija ima široku upotrebu u stručnoj literaturi kako u našoj zemlji, tako iu inostranstvu.
Određivanje vrste bakterija bez upotrebe umjetnih hranjivih podloga nemoguće je. S tim u vezi, može se primijetiti da pri uzgoju bakterija formiraju vrlo karakteristične kolonije. U ovom slučaju, jedan izgled se može koristiti za procjenu vrste bakterija.
Ako pronađete grešku, označite dio teksta i kliknite Ctrl+Enter.
1.5. Moderna klasifikacija bakterija U savremenoj taksonomiji bakterija razvila se situacija koja je karakteristična i za klasifikaciju drugih organizama: napravljen je napredak u stvaranju filogenetskog sistema klasifikacije koji odražava glavne pravce evolucijskog razvoja i odnos predstavnika bakterija. određene taksone, ali vještačke pojave zadržavaju svoj značaj - tipične klasifikacije, pogodnije za identifikaciju mikroorganizama. Trenutno ne postoji detaljan evolucijski sistem prokariota i, najvjerovatnije, rješenje ovog problema nije stvar bliske budućnosti. Osobine prokariota u oblasti morfološke, fiziološke, biohemijske i genetičke organizacije ukazuju na neprimjenjivost dobro razvijenih principa koji se koriste u izgradnji sistema viših organizama na njih. Ne zadržavajući se na historijskim aspektima problema taksonomije bakterija, treba napomenuti da je najprihvatljiviji sistem filogenetske klasifikacije za prokariote sistem zasnovan na podudaranju nukleotidne sekvence u 16S rRNA. Ovaj sistem činio je osnovu 2. izdanja višetomne enciklopedije prokariota - Bergeyjevog priručnika za sistematsku bakteriologiju (Bergey's Guide to Systematic Bacteria), čiji je prvi tom objavljen 2001. U ovom radu svi prokarioti su podijeljeni na 26 filogenetskih "grana" (grupa) na osnovu strukture njihove 16S-rRNA; 23 "grana" predstavljaju eubakterije, a tri arhebakterije. Treba naglasiti da veliki broj ovih filogenetskih grupa sadrži prokariotske vrste koje nisu izolovane kao čiste kulture i stoga još nisu detaljno proučavane. Za predstavnike ovih vrsta, trenutno su poznate samo 16S rRNA nukleotidne sekvence. Od 23 grupe eubakterija, dvije filogenetske grupe predstavljaju gram-pozitivne bakterije, preostale grupe su gram-negativne. Gram-negativne bakterije se sastoje od velike grupe proteobakterija (Proteobacteria) i 20 grupa drugih bakterija sa ovom vrstom ćelijskog zida. Kratak opis proteobakterija, kojima su mitohondriji i hloroplasti većine eukariota najbliži u sastavu 16S-rRNA, dat je u tabeli 1. 2. Proteobakterije su grupa gram-negativnih bakterija koja je vrlo heterogena u morfološkom, fiziološkom i biohemijskom smislu. Predstavnike ove grupe odlikuju svi vidovi energetskog metabolizma i ishrane. Ćelije većine vrsta Proteobacteria imaju štapićasti, sferični ili vibrioidni oblik; razmnožavaju se uglavnom binarnom fisijom, ali neke vrste karakteriziraju pupanje i formiranje plodišta u složenom ćelijskom ciklusu. Ova grupa sadrži i pokretne zbog flagele i nepokretne bakterije. S obzirom na molekularni kiseonik, proteobakterije se klasifikuju kao obavezni aerobi, obvezni i fakultativni anaerobi. Grupa Proteobacteria podijeljena je u pet podgrupa na osnovu razlika u 16S rRNA: alfa, beta, gama, delta i epsilon. Osim proteobakterija, gram-negativne eubakterije uključuju sljedeće glavne grupe eubakterija: termofili vodonika, zelene filamentne bakterije, zelene sumporne bakterije, cijanobakterije, spirohete, citofagi, bakteroidi, klamidije, planktomiceti, deinoofeksobacteri, deinoofeksobacteri, deinoofeksobacteri, itd. Filogenetske grupe gram-pozitivnih bakterija - Actinobacteria i Firmicutes. Grupu Actinobacteria (“grana aktinomiceta”) predstavljaju sljedeći rodovi bakterija sa visokim sadržajem GC parova u DNK: Geodermatophilus, Frankia, Streptomyces, Arthrobacter, Micrococcus, Actinomyces, Bifidobacterium, Propionibacterium, Actinoplanes, Corhodocobacterium, , Mycobacterium. Grupa Firmicutes („klostridijalna grana” - uglavnom gram-pozitivne bakterije sa niskim sadržajem GC-parova u DNK) se sastoji od sljedećih rodova: Clostridium, Lactococcus, Pediococcus, Streptococcus, Enterococcus, Leuconostoc, Listeria, Caryocophanoc, Staphylocophanoc, Staphylocophanoc , Sporosarcina, Bacillus, Desulfotomaculum, Heliobacterium, Mycoplasma, Ureaplasma, itd. Arhebakterije se dijele u tri filogenetske grupe: Crenarchaeota, Euryarchaeota i Korarchaeota. Grupu Crenarchaeota čine izuzetno termofilne bakterije, čiji većina predstavnika metabolizira sumpor, neki redukuju ione željeza i molibdena. Grupa Euryarchaeota uključuje obavezne anaerobne metanogene arhebakterije, kao i ekstremne termofile i halofile. Grupu Korarchaeota čine arhebakterije koje žive u vrelim sumpornim izvorima. Do danas nijedan od predstavnika ove grupe (koji posjeduju sličnu 16S rRNA) nije izolovan kao čista kultura, stoga njihove fenotipske karakteristike nisu dovoljno proučene. Završavajući razmatranje filogenetskih grana prokariota, treba napomenuti da predloženi filogenetski sistem, zasnovan na proučavanju nukleotidnih sekvenci samo jednog ribosomskog RNA gena, nije ništa drugo nego jedan od tehnički pogodnih i razvijenih sistema za naručivanje brojnih organizme kako bi ih identificirali, stoga je logično konstruirati ispravnu taksonomiju bakterija samo uzimajući u obzir ovu osobinu nije moguće. Najpoznatija i najkorišćenija fenotipska klasifikacija bakterija je ona predstavljena u devetom izdanju Burgeyjevog vodiča za bakterije. U ovom izdanju bakterije su podijeljene u četiri glavne kategorije (odjele) na osnovu strukture graničnog sloja ćelije: 1) Gracilicutes (od latinskog cutes - koža, gracilis - tanak) - gram-negativne eubakterije sa ćelijskim zidovima; 2) Firmicutes (od lat. firmus - jak) - gram-pozitivne eubakterije sa ćelijskim zidovima; 3) Tenericutes (od latinskog tener - mekan, nežan) - eubakterije, lišene ćelijskih zidova; 4) Mendosicutes (od latinskog mendosus - pogrešan) - arhebakterije, čiji se ćelijski zidovi razlikuju od sličnih struktura drugih prokariota. Odjel Gracilicutes uključuje bakterije različite morfologije sa gram-negativnim ćelijskim zidom. Reprodukcija se odvija uglavnom binarnom fisijom, neke bakterije se razmnožavaju pupanjem. Endospore se ne formiraju. Većina je pokretna: postoje sve vrste kretanja bakterija - uz pomoć flagela, klizanje, savijanje. Podjela uključuje aerobne, anaerobne i fakultativne anaerobne bakterije; fototrofne i hemotrofne bakterije. Odjeljenje je podijeljeno u tri klase: Scotobacteria, Oxyphotobacteria, Anoxyphotobacteria. Klasa Scotobacteria uključuje gram-negativne bakterije koje ne koriste svjetlosnu energiju u metaboličke svrhe, već je primaju samo kao rezultat redoks reakcija. Naziv klase dolazi od grčkog sotos - mrak. Ovo je najveća klasa bakterija. Klasa Anoksifotobakterija uključuje ljubičaste bakterije, zelene bakterije i heliobakterije, koje provode anoksigenu fotosintezu (bez oslobađanja molekularnog kisika). Klasu Oxyphotobacteria predstavljaju cijanobakterije i proklorofiti koji provode oksigensku fotosintezu (uz oslobađanje molekularnog kisika). Ova vrsta fotosinteze slična je fotosintezi koja se javlja u biljkama. Odjel Firmicutes uključuje bakterije s gram-pozitivnim ćelijskim zidom. Ćelije mogu imati različite oblike: štapiće, koke, filamentne, granaste. Neki predstavnici formiraju endospore. Većina njih je nepomična; pokretni oblici imaju peritrihozne flagele. Odjel obuhvata aerobne, anaerobne i fakultativno anaerobne bakterije. Odjel se sastoji od dvije klase: Firmibacteria, Thallobacteria. Klasa Firmibacteria uključuje veliki broj "ne-granajućih" Gram-pozitivnih bakterija. Klasa Thallobacteria uključuje bakterije čije ćelije su sposobne da se "grane". Odeljenje Tenerikute predstavljaju bakterije koje nemaju ćelijski zid. Zbog nepostojanja ćelijskog zida, oblik ćelija nije konstantan: u čistoj kulturi jedne vrste istovremeno su prisutne kokoidne, štapićaste, filamentne, kruškolike, diskaste i druge ćelije. Reprodukcija bakterija uključenih u ovaj odjeljak odvija se binarnom fisijom, pupanjem. Boja po Gramu je negativna. Karakteristično je formiranje malih kolonija koje rastu u agar. Mogu biti saprofitni, parazitski ili patogeni. Odjeljenje se sastoji od jedne klase Mollicutes (mikoplazme). Odeljenje Mendosicutes formiraju bakterije sa čvrstim ćelijskim zidom, ali bez murein peptidoglikana. Većina predstavnika su strogi anaerobi, od kojih mnogi imaju flagele. Vrste karakteriše ekološka i metabolička raznolikost, sposobnost da žive u ekstremnim uslovima. Odjeljenje se sastoji od jedne klase - Archaebacteria. U okviru četiri podjele (glavne kategorije) identificirano je 35 grupa (ili odjeljaka) bakterija, koje će u većoj ili manjoj mjeri biti okarakterisane u narednim poglavljima. Sljedeće grupe pripadaju odjelu Gracilicutes. Grupa 1. Spirohete. Grupa 2. Aerobne (ili mikroaerofilne), pokretne, spiralne (ili vibrioidne) Gram-negativne bakterije. Grupa 3. Nepokretne ili rijetko pokretne Gram-negativne zakrivljene bakterije. Grupa 4. Gram-negativne aerobne (ili mikroaerofilne) šipke i koke. Grupa 5. Opcioni aerobni gram-negativni štapići. Grupa 6. Gram-negativni anaerobni ravni, zakrivljeni ili spiralni štapići. Grupa 7. Bakterije koje vrše disimilaciju redukcije sumpora ili sulfata. Grupa 8. Anaerobne gram-negativne koke. Grupa 9. Rikecije i klamidija. Grupa 10. Anoksigene fototrofne bakterije. Grupa 11. Kiseoničke fototrofne bakterije. Grupa 12. Aerobne hemolitotrofne bakterije i srodni organizmi. Grupa 13. Bakterije koje pupe i (ili) stvaraju izrasline. Grupa 14. Bakterije sa ovojnicama. Grupa 15. Nefotosintetske klizne bakterije koje ne formiraju plodna tijela. Grupa 16. Klizne bakterije koje formiraju plodna tijela. Odeljenje Firmicutes obuhvata: Grupu 17. Gram-pozitivne koke. Grupa 18. Gram-pozitivni štapići i koke koje formiraju endospore. Grupa 19. Gram-pozitivni štapići pravilnog oblika, koji ne stvaraju spore. Grupa 20. Gram-pozitivni štapići nepravilnog oblika koji ne stvaraju spore. Grupa 21. Mycobacteria. Grupe 22–29. Actinomycetes. Odeljenju Tenericutes pripadaju: Grupa 30. Mikoplazme. Odjel Mendosicutes uključuje: Grupu 31. Metanogeni. Grupa 32. Sulfate-reducirajuće arhebakterije. Grupa 33. Ekstremno halofilne arhebakterije (halobakterije). Grupa 34. Arhebakterije koje nemaju ćelijski zid. Grupa 35. Ekstremno termofilne i hipertermofilne arhebakterije koje metaboliziraju sumpor. U zaključku, treba naglasiti da većinu mikroorganizama koji postoje u prirodnim zajednicama još uvijek treba izolirati u čiste kulture. Smatra se da se trenutno može uzgajati samo 0,1% ukupne mikrobne raznolikosti, a nije moguće uzgajati i identificirati preostale predstavnike bakterija, iako je oko 5 tisuća vrsta prokariota već izolirano i opisano u čistoj kulturi.