Mikroorganismu klasifikācija pēc to galvenajām formām. Baktērijas, to daudzveidība. Struktūra. Vitalitāte. Pamatjēdzieni: baktērijas
Baktēriju uzturs.
Uzturs.
pasīvā difūzija
Atvieglota difūzija
aktīvais transports
Pirmajā gadījumā barības vielu molekula veido kompleksu ar periplazmas telpas proteīnu, kas mijiedarbojas ar specifisku citoplazmas membrānas permeāzi. Pēc no enerģijas atkarīgas iekļūšanas caur citoplazmas membrānu komplekss "substrāts - periplazmatiskais proteīns - permeāze" disociējas ar substrāta molekulas izdalīšanos.
Aktīvās transportēšanas laikā ar transportējamās vielas ķīmisko modifikāciju notikumu ķēdē ietilpst: (1) membrānas enzīma-2 fosforilēšana no citoplazmas ar fosfoenolpiruvātu; (2) saistīšanās ar citoplazmas membrānas virsmu ar fosforilētu enzīma-2 substrāta molekulām; (3) no enerģijas atkarīga substrāta molekulas transportēšana citoplazmā; (4) fosfātu grupas pārnešana uz substrāta molekulu; (5) "substrāta-enzīma" kompleksa disociācija citoplazmā. Fosforilācijas dēļ substrāta molekulas uzkrājas šūnu citoplazmā un nespēj tās atstāt.
Baktēriju klasifikācija pēc pārtikas veida.
Barības vielu uzņemšanas ceļā baktērijas tiek klasificētas holofīti un Holozoja. Holofītu baktērijas (no grieķu val. holos- pilnīga un phyticos- kas saistīti ar augiem) nespēj izdalīt vidē enzīmus, kas noārda substrātus, kā rezultātā barības vielas patērē tikai izšķīdinātā, molekulārā veidā. Holozojas baktērijas (no grieķu val. holos- pilnīga un zoikos- saistīti ar dzīvniekiem), gluži pretēji, tiem ir eksoenzīmu komplekss, kas nodrošina ārēju uzturu - substrātu sadalīšanos molekulās, kas atrodas ārpus baktēriju šūnas. Pēc tam barības vielu molekulas nonāk holozoja baktēriju iekšpusē.
Pēc oglekļa avota izolēts no baktērijām autotrofi un heterotrofi. Autotrofi (no grieķu val. automašīnas- sevi, trofeju- pārtika) kā oglekļa avots tiek izmantots oglekļa dioksīds (CO 2), no kura tiek sintezētas visas oglekli saturošās vielas. Priekš heterotrofi (no grieķu valodas geteros - cits un trofeju- pārtika) oglekļa avoti ir dažādas organiskas vielas molekulārā formā (ogļhidrāti, daudzvērtīgie spirti, aminoskābes, taukskābes). Visaugstākā heterotrofijas pakāpe ir raksturīga prokariotiem, kas var dzīvot tikai citās dzīvās šūnās (piemēram, riketsijā un hlamīdijā).
Pēc slāpekļa avota prokariotus iedala 3 grupās: 1) slāpekli fiksējošās baktērijas (asimilē molekulāro slāpekli no atmosfēras gaisa); 2) baktērijas, kas patērē neorganisko slāpekli no amonija sāļiem, nitrītiem vai nitrātiem; 3) baktērijas, kas asimilē organiskajos savienojumos (aminoskābes, purīni, pirimidīni utt.) esošo slāpekli.
Pēc enerģijas avota baktērijas ir sadalītas fototrofi un ķīmijtrofi. fototrofās baktērijas , tāpat kā augi, spēj izmantot saules enerģiju. Fototrofiskie prokarioti neizraisa slimības cilvēkiem. Ķīmijtrofiskās baktērijas saņem enerģiju redoksreakcijās.
Pēc elektronu donoru būtības litotrofi(no grieķu val. litos- akmens) un organotrofi. Plkst litotrofi (chemolitotrofi ) neorganiskās vielas darbojas kā elektronu donori (H 2, H 2 S, NH 3, sērs, CO, Fe 2+ u.c.). Elektronu donori organotrofi (ķīmiskie organotrofi ) ir organiskie savienojumi – ogļhidrāti, aminoskābes u.c.
Lielākajai daļai cilvēku patogēno baktēriju ir ķīmijorganotrofs (ķīmoheterotrofs) uztura veids; chemolitotrophic (chemoautotrophic) tips ir retāk sastopams.
Ar spēju sintezēt organiskos savienojumusĶīmotrofās baktērijas tiek klasificētas prototrofi, auksotrofi un hipotrofi. prototrofās baktērijas sintezēt visas nepieciešamās organiskās vielas no glikozes un amonija sāļiem. Baktērijas sauc auksotrofi ja tie nespēj sintezēt kādu organisku vielu no norādītajiem savienojumiem. Ārkārtējo vielmaiņas aktivitātes zuduma pakāpi sauc par nepietiekamu uzturu. hipotrofiskas baktērijas nodrošina to vitālo aktivitāti, reorganizējot saimniekorganisma struktūras vai metabolītus.
Papildus ogleklim un slāpeklim, baktēriju pilnvērtīgai dzīvei ir nepieciešami sēra, fosfora un metālu joni. Sēra avoti ir aminoskābes (cisteīns, metionīns), vitamīni, kofaktori (biotīns, liposkābe utt.), sulfāti. Fosfora avoti ir nukleīnskābes, fosfolipīdi, fosfāti. Pietiekami augstā koncentrācijā baktērijām nepieciešams magnijs, kālijs, kalcijs, dzelzs; daudz mazākos daudzumos - cinks, mangāns, nātrijs, molibdēns, varš, niķelis, kobalts.
augšanas faktori– Tās ir vielas, kuras baktērijas pašas nespēj sintezēt, taču tām tās ir ļoti vajadzīgas. Aminoskābes, slāpekļa bāzes, vitamīni, taukskābes, dzelzs porfirīni un citi savienojumi var darboties kā augšanas faktori. Lai radītu optimālus apstākļus baktēriju dzīvībai svarīgai darbībai, barības vielu barotnēm jāpievieno augšanas faktori.
Metabolisms, enerģijas pārveide
A) Konstruktīva vielmaiņa.
Obligāta baktēriju uztura fāze ir barības vielu asimilācija, tas ir, to iekļaušana izmainītā vai modificētā veidā sintētiskās reakcijās šūnu komponentu un struktūru reprodukcijai.
Olbaltumvielu metabolisms baktērijās tas var notikt 3 fāzēs: primārā olbaltumvielu sadalīšanās, sekundārā olbaltumvielu sadalīšanās un olbaltumvielu sintēze. Olbaltumvielu molekulu primāro sadalīšanos līdz peptoniem veic eksoenzīmi - eksoproteāzes, ko baktērijas izdala vidē. Sekundārā sabrukšana notiek endoenzīmu (endoproteāžu) ietekmē, kas piemīt visām baktērijām. Šis process notiek baktēriju šūnā un sastāv no peptīdu sadalīšanās to sastāvā esošajās aminoskābēs. Pēdējos var izmantot nemainītā veidā vai pakļaut ķīmiskām pārvērtībām (deaminēšana, dekarboksilēšana utt.), kuru rezultātā parādās amonjaks, indols, sērūdeņradis, keto skābes, spirts, oglekļa dioksīds uc Šo savienojumu noteikšana. ir diagnostiska nozīme bakterioloģijā.
Līdz ar olbaltumvielu šķelšanās reakcijām notiek arī to sintēzes reakcijas. Dažas baktērijas veido olbaltumvielas no gatavām aminoskābēm, kas iegūtas ārējā uztura rezultātā, bet citas baktērijas neatkarīgi sintezē aminoskābes no vienkāršiem savienojumiem, kas satur slāpekli un oglekli. Aminoskābju sintēzi var veikt aminēšanas, transaminēšanas, amidēšanas, karboksilēšanas reakcijās. Lielākā daļa prokariotu spēj sintezēt visas aminoskābes, kas veido šūnu proteīnus. Aminoskābju biosintēzes iezīme ir kopīgu biosintēzes ceļu izmantošana: trikarbonskābes cikls, glikolīze, oksidatīvā pentozes-fosfāta ceļš utt. Galvenais aminoskābju sintēzes savienojums ir piruvāts un fumarāts.
ogļhidrātu metabolisms tas atšķiras ar autotrofiem un heterotrofiem (1. shēma). Autotrofās baktērijas no oglekļa dioksīda sintezē visus nepieciešamos ogļhidrātus. Izejvielas ogļhidrātu veidošanai heterotrofajās baktērijās var būt: (1) viena, divu un trīs oglekļa savienojumi; un (2) polisaharīdi (ciete, glikogēns, celuloze). Lai sadalītu pēdējo, daudzām heterotrofajām baktērijām ir eksoenzīmi (amilāze, pektināze u.c.), kas hidrolizē polisaharīdus, veidojot glikozi, maltozi, fruktozi utt.
Autotrofajās baktērijās Kalvina ciklā no oglekļa dioksīda veidojas ribulozes fosfāts-fosfors-glicerīnskābe, kas tiek iekļauta glikolīzes reakcijās, kas notiek pretējā virzienā. Reversās sintēzes galaprodukts ir glikoze.
Heterotrofās baktērijas veido glikozi no viena, divu un trīs oglekļa savienojumiem, iekļaujot tos arī reversās glikolīzes reakcijā. Sakarā ar to, ka dažas glikolīzes reakcijas ir neatgriezeniskas, heterotrofi ir izveidojuši īpašas fermentatīvas reakcijas, kas ļauj apiet kataboliskā ceļa neatgriezeniskās reakcijas.
Kad polisaharīdus šķeļ heterotrofās baktērijas, iegūtie disaharīdi nonāk šūnās un maltozes, saharozes un laktozes ietekmē tiek hidrolizēti un sadalās monosaharīdos, kas pēc tam tiek fermentēti vai iekļauti cukura savstarpējās konversijas reakcijās.
lipīdu metabolisms. Gan eksogēni lipīdi, gan intersticiāla metabolisma amfibolīti var kalpot par izejmateriāliem lipīdu veidošanai baktērijās. Eksogēni lipīdi ir pakļauti baktēriju lipāzēm un citiem lipolītiskiem enzīmiem. Daudzu veidu baktērijas absorbē glicerīnu, kas kalpo kā plastmasas materiāla un enerģijas avots. Endogēni avoti lipīdu sintēzei var būt acetilkoenzīms A, propionil-APB, malonil-APB (ACP - acetyl-transporting protein), fosfodioksiacetons utt.
Sākotnējais substrāts taukskābju sintēzei ar pāra oglekļa atomu skaitu ir acetilkoenzīms A, taukskābēm ar nepāra oglekļa atomu skaitu propionil-APB un malonil-APB. Divkāršo saišu veidošanās skābes molekulā aerobos prokariotos notiek, piedaloties molekulārajam skābeklim un fermentam desaturāzei. Anaerobos prokariotos divkāršās saites tiek ieviestas sintēzes sākumā dehidratācijas reakcijas rezultātā. Sākotnējais fosfolipīdu sintēzes substrāts ir fosfodioksiacetons (glikolītiskā ceļa starpsavienojums), kura reducēšanās rezultātā veidojas 3-fosfoglicerīns. Pēc tam pēdējam pievieno 2 taukskābju atlikumus kompleksa veidā ar APB. Reakcijas produkts ir fosfatidīnskābe, kuras aktivizēšana ar CTP un sekojoša pievienošana serīna, inozitola, glicerīna vai cita savienojuma fosfātu grupai noved pie atbilstošo fosfolipīdu sintēzes.
Mikroorganismi, kas ir auksotrofi un hipotrofiski taukskābēm (piemēram, mikoplazmas), iegūst tos gatavus no saimniekšūnām vai barotnes.
Mononukleotīdu apmaiņa. Purīna un pirimidīna mononukleotīdi ir būtiskas DNS un RNS sastāvdaļas. Daudzi prokarioti spēj gan izmantot jau gatavas purīna un pirimidīna bāzes, to nukleozīdus un nukleotīdus, kas atrodas uzturvielu barotnē, gan sintezēt tos no zemas molekulmasas vielām. Baktērijās ir fermenti, kas katalizē šādus eksogēno purīna un pirimidīna atvasinājumu savstarpējās konversijas posmus: slāpekļa bāze - nukleozīds - nukleotīds (mono - di - - trifosfāts).
Purīna un pirimidīna mononukleotīdu sintēze de novo veic neatkarīgos veidos. Purīna nukleotīdu sintēzes laikā secīgu enzīmu reakciju rezultātā veidojas inozīnskābe, no kuras, ķīmiski modificējot purīna gredzenu, tiek sintezētas adenilskābes (AMP) un guanilskābes (GMP). Pirimidīna nukleotīdu sintēze sākas ar orotidilskābes veidošanos, kuras dekarboksilēšana dod uridilskābi (UMP). No pēdējās veidojas UTP, kuras acilēšana noved pie CTP veidošanās.
Dezoksiribonukleotīdi veidojas atbilstošo ribonukleotīdu reducēšanās rezultātā difosfātu vai trifosfātu līmenī. DNS specifiskā nukleotīda, timidilskābes, sintēze notiek dezoksiuridilskābes fermentatīvās metilēšanas ceļā.
Jonu apmaiņa. Minerālie savienojumi - joni, NH 3 +, K +, Mg 2+, Fe 2+, SO 4 2-, PO 4 3- un citas baktērijas tiek iegūtas no vides gan brīvā, gan ar citām organiskām asociētā stāvoklī. vielas. Katjoni un anjoni tiek transportēti baktēriju šūnā dažādos veidos, kas aprakstīti § 3. Jonu iekļūšanas ātrumu baktērijas šūnā ietekmē barotnes pH un pašu mikroorganismu fizioloģiskā aktivitāte.
B) Baktēriju elpošana (enerģijas metabolisms).
Visi dzīvības procesi ir atkarīgi no enerģijas, tāpēc enerģijas iegūšana ir ārkārtīgi svarīgs prokariotu metabolisma aspekts. Viņi iegūst enerģiju no anaerobās un aerobās elpošanas.
Elpa, vai bioloģiskā oksidācija ir katabolisks elektronu pārneses process no donorvielas uz akceptorvielu, ko pavada enerģijas uzkrāšanās makroerģiskajos savienojumos. . Elpošana notiek katabolisko reakciju procesā, kā rezultātā sarežģītas organiskās vielas, sadaloties, izdala enerģiju un pārvēršas vienkāršos savienojumos. Makroerģiskajās vielās (ATP, GTP, UTP u.c.) uzkrātā enerģija tiek izmantota anaboliskajās reakcijās.
Atbilstoši elpošanas veidam mikroorganismus iedala obligātie (stingri) aerobi, obligātie anaerobi un fakultatīvie anaerobi.
obligātie aerobi nepieciešams brīvs skābeklis. Organiskie savienojumi (ogļhidrāti, tauki, olbaltumvielas) ir elektronu donori cilvēka patogēnos aerobos-ķīmoorganotrofos, un molekulārais skābeklis ir elektronu akceptors. Enerģijas uzkrāšanās ATP formā ķīmijorganotrofiskajos aerobos notiek elektronu donoru oksidatīvās fosforilēšanas laikā. Aerobos ir citohromi (piedalās elektronu pārnesē), kā arī fermenti (katalāze, superoksīda dismutāze, peroksidāze), kas inaktivē toksiskos skābekļa radikāļus, kas rodas elpošanas laikā. Superoksīda dismutāze inaktivē toksiskāko metabolītu, superoksīda radikāli H 2 O 2 . Enzīms katalāze pārvērš H 2 O 2 par H 2 O un O 2.
Īpaša aerobu grupa ir mikroaerofilās baktērijas, kas, lai gan tiem ir nepieciešams skābeklis enerģijai, labāk aug ar paaugstinātu CO 2 saturu, piemēram, ģints baktērijas Kampilobaktērijas un Helicobacter.
obligātie anaerobi nav nepieciešams brīvais skābeklis, gluži pretēji, pat nelielos daudzumos skābeklis uz tiem iedarbojas toksiski. Elektronu donori cilvēka patogēnos anaerobos-ķīmoorganotrofos ir dažādi organiskie savienojumi (galvenokārt ogļhidrāti). Elektronu akceptors ķīmijorganotrofajos anaerobos ir organiski skābekli saturoši savienojumi - skābes vai ketoni, tas ir, elektronu akceptors ir skābeklis, kas saistīts ar organisko fragmentu. Enerģijas uzglabāšana šajos prokariotos notiek substrāta fosforilēšanās laikā. Obligātajiem anaerobiem, kā likums, nav citohromu un enzīmu, kas inaktivē skābekļa radikāļus (katalāzes un superoksīda dismutāzes negatīvi).
Ķīmolitotrofiskajos anaerobos, kas nav patogēni cilvēkiem, elektronu akceptors ir neorganiskie skābekli saturoši savienojumi - nitrāti, sulfāti, karbonāti.
Īpaša anaerobu grupa ir aerotolerantas baktērijas, kas spēj vairoties atmosfēras skābekļa klātbūtnē, bet neizmanto to kā elektronu akceptoru (piemēram, pienskābes baktērijas). Aerotoleranti katalāze un superoksīda dismutāzes prokarioti ir pozitīvi.
Fakultatīvie anaerobi spēj pastāvēt gan skābekļa, gan bezskābekļa vidē. To elektronu donori ir organiskas vielas; elektronu akceptori, atkarībā no vides apstākļiem, ir molekulāri vai skābeklis, kas saistīts ar organiskiem un neorganiskiem savienojumiem. Fakultatīvie anaerobi var uzkrāt enerģiju gan oksidatīvās, gan substrāta fosforilēšanas laikā. Tāpat kā aerobos, šai baktēriju grupai ir citohromi un antioksidantu aizsardzības enzīmi.
Galvenais substrāts enerģijas iegūšanai ir ogļhidrāti, kas dažādu elpošanas veidu ķīmiskoheterotrofajos prokariotos var tikt katabolizēti par acetilkoenzīmu A (“aktivētā etiķskābe”). Lipīdi un proteīni var darboties kā enerģijas substrāti, jo acetilkoenzīms A ir arī viens no to metabolisma starpproduktiem (2. shēma).
Ogļhidrātu katabolisms ķīmijorganotrofiskajos prokariotos ietver: (a) anaerobos procesus — glikolīzi, pentozes fosfāta ceļu un ketodeoksifosfoglikonāta ceļu; b) aerobais process – trikarbonskābes cikls (Krebsa cikls). Anaerobie procesi notiek visos prokariotos, savukārt aerobais process ir raksturīgs tikai obligātajiem aerobiem un fakultatīvajiem anaerobiem. Anaerobo ceļu pamatā ir substrāta fosforilēšana, savukārt aerobos procesu pamatā ir oksidatīvā fosforilēšana.
Jēdzienu definīcija.
Sterilizācija, dezinfekcija un antiseptiķi ir mūsdienu medicīnas un īpaši ķirurģiskās prakses neatņemama sastāvdaļa. Izpratne par šo metožu principiem un praktisko pielietojumu ir būtiska, jo daudzi potenciāli patogēni mikroorganismi var izdzīvot ārpus saimniekorganisma ilgu laiku, tiem ir augsta izturība pret fizikāliem un ķīmiskiem dezinfekcijas līdzekļiem un salīdzinoši viegli izplatīties no cilvēka uz cilvēku.
Antiseptiķi- patogēnu vai oportūnistisku mikroorganismu iznīcināšana vai augšanas novēršana ar ķīmiskām metodēm. Šo terminu parasti lieto, lai apzīmētu ķīmiska preparāta ārēju lietošanu dzīviem audiem.
antiseptisks- viela, kas kavē mikroorganismu augšanu vai iznīcina (neiedarbojoties uz baktēriju sporām). Šis termins attiecas uz vielām, kuras lieto lokālai iedarbībai uz dzīviem audiem.
Aseptika nozīmē sepses neesamību, bet kopumā šis termins tiek lietots, lai uzsvērtu dzīvu organismu neesamību. Aseptiskas metodes ir jebkura procedūra, kas paredzēta dzīvo organismu likvidēšanai un to atkārtotas inficēšanās novēršanai. Mūsdienu ķirurģiskās un mikrobioloģiskās metodes balstās uz aseptiskām procedūrām.
Biocīds- viela, kas iznīcina visus dzīvos mikroorganismus, gan patogēnos, gan nepatogēnos, tostarp sporas.
Biostat- līdzeklis, kas novērš mikroorganismu augšanu, bet ne vienmēr tos nogalina.
Dekontaminācija- mikroorganismu noņemšana bez kvantitatīvās noteikšanas. Šis termins ir relatīvs; galīgo mikrobu izņemšanu var veikt ar sterilizāciju vai dezinfekciju.
Dezinfekcija- process, kas samazina vai likvidē visus patogēnus, izņemot sporas.
Baktērijas iznīcinošs līdzeklis- viela, kas iznīcina mikroorganismus, īpaši patogēnus. Baktērijas iznīcināšanas līdzeklis neiznīcina sporas.
Sanācija- metode, ar kuras palīdzību mikrobu piesārņojums tiek samazināts līdz “drošam” līmenim. Šī metode iepriekš tika izmantota nedzīvu objektu "attīrīšanai".
Sterilizācija- fizikālo faktoru un (vai) ķīmisko vielu izmantošana visu veidu mikrobu dzīvības pilnīgai iznīcināšanai vai iznīcināšanai.
Sterilizācija.
Sterilizāciju definē kā visu mikroorganismu un to sporu iznīcināšanu vai izņemšanu (filtrējot). Sterilizāciju parasti veic, izmantojot siltumu. Sterilizācija, kas ir viena no ikdienas gaitām mikrobioloģijas laboratorijas darbā, ir būtiska metode, lai nodrošinātu, ka kultūras, iekārtas, trauki un barotnes atbalsta tikai nepieciešamo mikroorganismu augšanu, bet pārējie mikrobi tiek iznīcināti. Ir šādi sterilizācijas veidi: kalcinēšana degļa liesmā, vārīšana, darbība ar plūstošu tvaiku, tvaiks zem spiediena autoklāvā, sausais karstums, pasterizācija, tindalizācija, ķīmiskā, aukstā (mehāniskā) sterilizācija.
Sterilizācijas metožu izvēle.
Izvēloties sterilizācijas metodes, jāņem vērā šādas prasības:
1. Aktivitāte: baktericīda, sporicīda, tuberkulocīda, fungicīda un virocīda.
2. Procedūras ātrums: sterilizācija jāveic pēc iespējas ātrāk.
3. Caurlaidība: Sterilizācijas līdzekļiem jāspēj iekļūt caur iepakojumu un instrumenta iekšpusē.
4. Saderība: nedrīkst būt izmaiņas to materiālu struktūrā vai funkcijās, kas tiek sterilizēti vairākas reizes.
5. Netoksicitāte: nedrīkst radīt draudus cilvēku veselībai un videi.
6. Organisko materiālu noturība: organisko materiālu klātbūtnē nedrīkst samazināt sterilizācijas efektivitāti.
7. Pielāgojamība: iespēja izmantot lieliem un maziem sterilizēta materiāla apjomiem.
8. Kontrole laika gaitā: apstrādes ciklam jābūt viegli un precīzi kontrolētam.
9. Cena: saprātīgas aprīkojuma, uzstādīšanas un ekspluatācijas izmaksas.
 Fiziskie sterilizatori
Fiziskie sterilizatori
slapjš sirsnīgi, kas veidojas tvaika autoklāvēšanas procesā, ir galvenais sterilizējošais līdzeklis, ko izmanto klīniskās mikrobioloģijas laboratorijās. Autoklāvus izmanto barotņu, ugunsizturīgo materiālu sterilizēšanai un infekciozo atkritumu apstrādei. Tvaika sterilizators jeb autoklāvs ir izolēta spiediena kamera, kas izmanto piesātinātu tvaiku, lai radītu augstu temperatūru (1. attēls). Gaiss tiek izņemts no kameras ar masas pārvietošanu vai vakuumu. Visbiežāk izmantotie autoklāvi ar aizstāšanu pēc svara. Kamerā tiek ievadīts vieglāks tvaiks, lai izspiestu smagāku gaisu. Īslaicīga saspiesta tvaika iedarbība var iznīcināt baktēriju sporas. Parastā barotņu un citu materiālu sterilizācijai ekspozīcijas laiks ir 15 minūtes 121°C temperatūrā un 1,5 kg spiedienā uz 1 kvadrātcentimetru. Infekciozo atkritumu gadījumā ekspozīcijas laiks tiek palielināts līdz 30-60 minūtēm. Papildus pareizajam laikam un temperatūrai sterilizācijai ļoti svarīgs ir tiešs kontakts ar tvaiku. Rīkojoties ar infekciozu materiālu, jānodrošina maksimāla tvaika iekļūšana atkritumos. Šāds materiāls jāapstrādā 132ºС temperatūrā. Pretaudzēju preparātus, toksiskas ķīmiskas vielas un radioizotopus, kas var nesadalīties, un nestabilas ķīmiskas vielas nedrīkst lietot autoklāvā, jo tās var iztvaikot un izplatīties pa kameru, ja tās tiek pakļautas karstumam.
 Sausā karstuma sterilizācija izmanto materiāliem, kurus nevar sterilizēt ar tvaiku, jo var tikt bojāts vai materiāls nav tvaiku necaurlaidīgs. Sausais karstums ir mazāk efektīvs nekā mitrs, un tam ir nepieciešams ilgāks iedarbības laiks un augstāka temperatūra. Sterilizāciju ar sausu karstumu parasti veic sausā siltuma skapī (2. att.). Sausā karstuma sterilizācijas mehānisms ir oksidatīvs process. Materiālu piemēri, kuriem tiek izmantota sausā karstuma sterilizācija, ir eļļas, pulveri, asi instrumenti un stikla trauki. Sausais karstums vai termiskā inaktivācija-sterilizācija tiek izmantota kā alternatīvas metodes infekciozo atkritumu apstrādei.
Sausā karstuma sterilizācija izmanto materiāliem, kurus nevar sterilizēt ar tvaiku, jo var tikt bojāts vai materiāls nav tvaiku necaurlaidīgs. Sausais karstums ir mazāk efektīvs nekā mitrs, un tam ir nepieciešams ilgāks iedarbības laiks un augstāka temperatūra. Sterilizāciju ar sausu karstumu parasti veic sausā siltuma skapī (2. att.). Sausā karstuma sterilizācijas mehānisms ir oksidatīvs process. Materiālu piemēri, kuriem tiek izmantota sausā karstuma sterilizācija, ir eļļas, pulveri, asi instrumenti un stikla trauki. Sausais karstums vai termiskā inaktivācija-sterilizācija tiek izmantota kā alternatīvas metodes infekciozo atkritumu apstrādei.
Pasterizācija iznīcina patogēnus, ātri uzsildot vielu līdz 71,1°C 15 sekundes, kam seko ātra dzesēšana. Pasterizācija nav sterilizācija, jo ne visi mikroorganismi ir pret to jutīgi. Šī metode novērsa tādu slimību pārnešanu kā kuņģa-zarnu trakta tuberkuloze un Q drudzis.
Tindalizācija ir intermitējoša termiskās sterilizācijas metode, ko var izmantot, lai iznīcinātu visas šķīdumos esošās baktērijas. Tā kā augošās baktērijas viegli nogalina īsas vārīšanās rezultātā (5 reizes 1 stundas laikā pa 5 minūtēm), viss, kas jādara, ir jāļauj šķīdumam noteiktu laiku nostāvēties, pirms karstums izjauc sporu nobriešanu. ievērojams siltuma pretestības zudums.
Filtrēšana ir process, ko izmanto, lai noņemtu mikrobus un mikroskopiskas daļiņas no šķīdumiem, gaisa un citām gāzēm. Laboratorijā visbiežāk izmanto sterilizāciju ar filtrēšanu, lai apstrādātu diagnostiku, barotnes, audu barotnes, serumus, šķīdumus, kas satur seruma komponentus. Vēl viens izplatīts filtrēšanas pielietojums ir gaisa un gāzu sterilizācija. Plastmasas vai papīra membrānfiltri, kas izceļas ar poru diametru (apmēram no 12 līdz 0,22 µm) un tiek izmantoti mehāniskai atdalīšanai, kalpo arī mikrobu savākšanai no šķidrumiem mikroskopiskai izmeklēšanai vai kultivēšanai tieši uz filtra, kad tos novieto uz virsmas, kas samērcēta ar barības vielām. vidējs.
ultravioletais starojums ir elektromagnētisko viļņu starojuma veids, kas iedarbojas uz šūnu nukleīnskābi. Mikroorganismi ir ļoti jutīgi pret ultravioleto staru iedarbību ar viļņa garumu 254 nm. Ultravioleto gaismu visplašāk izmanto, lai iznīcinātu mikroorganismus gaisā vai uz virsmām. Citi lietojumi ir noteiktu ķīmisko vielu un plastmasas aukstā sterilizācija farmācijas vajadzībām, seruma sterilizācija šūnu kultūrām un ūdens dezinfekcija. Būtisks ultravioletā starojuma kā sterilizatora trūkums ir tā nespēja iekļūt materiālos.
jonizējošā radiācija elektromagnētiskajā spektrā tam ir letāla ietekme uz mikroorganismiem. Šis spektrs ietver mikroviļņus, γ-starus, rentgenstarus un elektronu plūsmu. Jonizējošā starojuma letālā iedarbība rodas tiešas iedarbības dēļ uz mērķa molekulu, kā rezultātā enerģija tiek pārnesta uz molekulu; un netiešās darbības dēļ - radikāļu difūzija.
ultraskaņas enerģija ar zemu frekvenci inaktivē mikroorganismus ūdens šķīdumos. Ultraskaņas apstrādes fiziskā ietekme rodas kavitācijas dēļ. Ultraskaņas tīrītājus un citas ierīces bieži izmanto instrumentu tīrīšanai, taču tās neuzskata par sterilizatoriem. Tomēr ultraskaņas kombinācija ar ķīmisko apstrādi nogalina mikroorganismus.
Ķīmiskie sterilizatori
2 % glutaraldehīds Kā šķidrs ķīmiskais sterilizators to iepriekš plaši izmantoja tādu medicīnisku un ķirurģisku materiālu apstrādei, kurus nevar sterilizēt ar karsēšanu vai apstarošanu. Glutaraldehīdu izmanto arī vakcīnu gatavošanā.
Dezinfekcija.
Dezinfekciju var veikt ar ķīmiskām metodēm vai vārot. Vārīšana ir efektīva instrumentu, piemēram, adatu un šļirču, dezinfekcijas metode, ja autoklāvs nav pieejams. Iepriekš iztīrītus medicīniskos instrumentus vajadzētu vārīt 20 minūtes. Ķīmisko dezinfekciju izmanto karstumjutīgām iekārtām, kuras var sabojāt augsta temperatūra. Plaši tiek izmantoti ķīmiskie dezinfekcijas līdzekļi, piemēram, hlora komponenti, etilspirts un izopropilspirts, ceturtā amonija komponenti un glutaraldehīds.
Ķīmiskie dezinfekcijas līdzekļi.
Alkohols (etil un izopropil), izšķīdināts ūdenī līdz 60-85% koncentrācijai, ir ļoti efektīvs dezinfekcijā. Alkoholi ir baktericīdi, fungicīdi un tuberkulocīdi, bet neietekmē sporas. Etilspirtam ir plašāks virocīdās aktivitātes spektrs nekā izopropilspirtam, tāpēc tas ir efektīvāks pret lipofīliem un hidrofiliem vīrusiem.
risinājums 37% formaldehīds, ko sauc par formalīnu, var izmantot kā sterilizatoru, savukārt tā koncentrāciju 3-8% var izmantot kā dezinfekcijas līdzekli.
Fenols tīrā veidā netiek izmantots kā dezinfekcijas līdzeklis, jo tas ir toksisks, spēj izraisīt audzēju attīstību un koroziju. Fenola atvasinājumi, kuros funkcionālā grupa (hlors, broms, alkilgrupa, benzils, fenils, amils) aizstāj vienu no ūdeņraža atomiem aromātiskajā gredzenā, tiek plaši izmantoti kā dezinfekcijas līdzekļi. Šī aizstāšana samazina fenola trūkumus. Fenola sastāvdaļas iznīcina mikrobus, jo tiek inaktivētas enzīmu sistēmas, izgulsnējas olbaltumvielas un tiek bojāta šūnu siena un membrāna. Parasti izmanto 2-5% koncentrāciju, zemākām koncentrācijām nepieciešama ilgāka iedarbība.
Halogēni. Laboratorijas praksē dezinfekcijai izmanto tikai hloru un jodu. Sakarā ar to, ka hlors ir spēcīgs oksidētājs, tiek uzskatīts, ka tas oksidējot iznīcina mikrobus. Tiek uzskatīts, ka jods nogalina mikroorganismus, reaģējot ar aminoskābju N-H un S-H grupām, kā arī ar aminoskābes tirozīna fenola grupu un nepiesātināto taukskābju oglekļa-oglekļa dubultsaitēm. Tradicionālā apstrāde ietver 2-5% formaldehīda šķīduma izsmidzināšanu tvaika klātbūtnē 60-80ºC temperatūrā.
Antiseptiķi.
Antiseptiskus līdzekļus var atrast mikrobioloģiskajās laboratorijās, galvenokārt vielās, ko izmanto roku mazgāšanai. Gadījumos, kad ārstniecības personas sniedz neatliekamo palīdzību pacientiem, izmantojot antibakteriālus līdzekļus saturošas vielas, tas samazina slimnīcu infekciju skaitu. Roku mazgāšanas līdzekļos visbiežāk sastopamās ķīmiskās vielas ir spirti, hlorheksidīna glikonāts, jodofori, hloroksilenols un triklozāns.
Tradicionālās atkritumu un atkritumu apstrādes metodes ir sadedzināšana un sterilizācija ar tvaiku.
Degšana ir izvēles metode atkritumu un gružu apstrādei. Šī metode padara atkritumus neinfekciozus, kā arī maina to formu un izmēru. Sterilizācija ir efektīva atkritumu apstrādes metode, taču tā nemaina tā formu. Sterilizācija ar tvaiku autoklāvā 121°C temperatūrā vismaz 15 minūtes iznīcina visa veida mikrobu dzīvības, tostarp lielu skaitu baktēriju sporu. Šāda veida pilnīgu sterilizāciju var veikt arī, izmantojot sausu karstumu 160-170ºC 2-4 stundas. Tomēr ir jānodrošina, lai sauss karstums saskartos ar sterilizējamo materiālu. Tāpēc pudeles, kurās ir šķidrums, ir brīvi jāaizver ar korķiem vai vates tamponiem, lai tvaiks un siltums varētu apmainīties ar pudelēs esošo gaisu. Bioloģiski bīstamās tvertnes, kurās ir atkritumi, ir cieši jāsasien. Sterilizēts bioloģiski bīstams materiāls jānoslēdz atbilstošos marķētos konteineros.
Sterilizācija ar tvaiku (autoklāvs). Infekciozie atkritumi tiek uzskatīti par dekontaminētiem, ja veģetatīvo baktēriju, sēnīšu, mikobaktēriju un lipīdus saturošo vīrusu skaits samazinās par 6 lg reizes, bet baktēriju endosporu skaits - par 4 lg reizes.
Baktēriju uzturs.
Uzturs. Baktēriju šūnas uzturā ir jāsaprot plastmasas materiāla un enerģijas absorbcijas un asimilācijas process transformatīvo reakciju rezultātā. . Prokariotu uztura veidi ir sarežģīti un daudzveidīgi. Tās atšķiras atkarībā no barības vielu iekļūšanas veida baktēriju šūnā, oglekļa un slāpekļa avotiem, enerģijas iegūšanas veida un elektronu donoru rakstura.
barības vielu transportēšana šūnā var veikt ar 3 mehānismiem: pasīvā difūzija, atvieglota difūzija un aktīvā transportēšana.
pasīvā difūzija ir nespecifisks no enerģijas atkarīgs process, kas tiek veikts pa vielu koncentrācijas gradientu (viela no vides ar lielāku koncentrāciju pasīvi, saskaņā ar osmozes likumiem, nonāk vidē ar mazāku koncentrāciju). Ar pasīvo difūziju baktēriju šūnā nonāk ierobežots vielu daudzums, daži joni, monosaharīdi. Vielu pārneses ātrums pasīvās difūzijas laikā ir nenozīmīgs un ir atkarīgs no transportējamo molekulu lipofilitātes un lieluma.
Atvieglota difūzija ir negaistošs vielu transports pa koncentrācijas gradientu ar permeāzes enzīmu palīdzību. Permeāzes ir specifiski membrānas proteīni, kas atvieglo vielu pāreju caur citoplazmas membrānu. Permeāze piestiprina pie sevis pārnestās vielas molekulu, kopā ar kuru tā pārvar citoplazmas membrānu, pēc tam "vielas-permeāzes" komplekss disociējas. Atbrīvotā permeāze tiek izmantota citu molekulu vadīšanai. Prokariotos tikai glicerīns iekļūst šūnā ar atvieglotu difūziju. Šajā gadījumā glicerīna intracelulārā koncentrācija atbilst tai, kas atrodas ārpus šūnas. Atvieglināta difūzija ir visraksturīgākā eikariotu mikroorganismiem.
aktīvais transports- tā ir no enerģijas atkarīga vielu pārnešana šūnā pret koncentrācijas gradientu ar specifisku enzīmu palīdzību. Absolūtais vairums vielu (joni, ogļhidrāti, aminoskābes, lipīdi u.c.) nokļūst baktēriju šūnā ar aktīvu transportu. Aktīvā transportēšana var notikt: (1) bez transportējamās vielas ķīmiskās modifikācijas; (2) ar ķīmisku modifikāciju.
Baktērijas ir šūnu struktūras prokariotiski mikroorganismi. To izmēri ir no 0,1 līdz 30 mikroniem. Mikrobi ir ārkārtīgi izplatīti. Viņi dzīvo augsnē, gaisā, ūdenī, sniegā un pat karstajos avotos, uz dzīvnieku ķermeņa, kā arī dzīvo organismu iekšpusē, tostarp cilvēka ķermenī.
Baktēriju sadalījums pa sugām balstās uz vairākiem kritērijiem, starp kuriem visbiežāk tiek ņemta vērā mikroorganismu forma un to telpiskā izplatība. Tātad, atkarībā no šūnu formas, baktērijas tiek sadalītas:
Coci - mikro-, diplo-, strepto-, stafilokoki, kā arī sarkīni;
Stieņa formas - monobaktērijas, diplobaktērijas un streptobaktērijas;
Vītņotās sugas - vibrios un spirohetas.
Beržeja determinants sistematizē visas zināmās baktērijas pēc to baktēriju identifikācijas principiem, kuras atradušas visplašāko izplatību praktiskajā bakterioloģijā, pamatojoties uz šūnu sienas struktūras atšķirībām un attiecībā pret Grama krāsojumu. Baktēriju apraksts dots pa grupām (sadalām), kas ietver dzimtas, ģintis un sugas; dažos gadījumos grupās ietilpst nodarbības un ordeņi. Cilvēkam patogēnās baktērijas ir iekļautas nelielā skaitā grupu.
Atslēga izšķir četras galvenās baktēriju kategorijas -
Gracillicutes [no lat. gracilis, graciozs, tievs, + cutis, āda] - suga ar plānu šūnu sieniņu, krāsota gramnegatīvs;
firmicutes [no lat. flrmus, stiprs, + cutis, āda] - baktērijas ar biezu šūnu sieniņu, krāsojums grampozitīvs;
Tenericutes [no lat. maigs, maigs, + cutis, āda] - baktērijām, kurām trūkst šūnu sienas(mikoplazmas un citi Mollicutes klases pārstāvji)
Mendosikūti [no lat. mendosus, neregulāras, + cutis, āda] - arhebaktērijas (metānu un sulfātu reducējošās, halofīlās, termofīlās un arhebaktērijas, bez šūnas sienas).
2. grupas Burgey noteicējs. Aerobās un mikroaerofīlās kustīgās vītņotas un izliektas gramnegatīvas baktērijas. Cilvēkam patogēnās sugas ir iekļautas Campylobacter, Helicobacters Spirillum dzimtās.
Beržeja noteicēja 3. grupa. Nekustīgas (retāk kustīgas) gramnegatīvas baktērijas. Nesatur patogēnas sugas.
Burgey noteicēja 4. grupa. Gramnegatīvi aerobās un mikroaerofīlās nūjiņas un koki. Cilvēkam patogēnās sugas iekļautas Legionellaceae, Neisseriaceae un Pseudomonada-ceae dzimtās, grupā ietilpst arī Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Franceisella, Kingella un Moraxella ģints patogēnās un oportūnistiskās baktērijas.
Beržeja noteicēja 5. grupa. Fakultatīvi anaerobie gramnegatīvi stieņi. Grupu veido trīs dzimtas - Enterobacteriaceae, Vibrionaceae un Pasteurellaceae, no kurām katra ietver patogēnās sugas, kā arī Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella un Streptobacillus ģinšu patogēnās un oportūnistiskās baktērijas.
Beržeja noteicēja 6. grupa. Gramnegatīvas anaerobas taisnas, izliektas un spirālveida baktērijas. Patogēnās un oportūnistiskās sugas ir iekļautas Bacteroides, Fusobacterium, Porphoromonas un Prevotelta ģintīs.
Beržeja noteicēja 7. grupa. Baktērijas, kas veic sulfāta vai sēra disimilācijas reducēšanu Neietver patogēnās sugas.
Beržeja noteicēja 8. grupa. Anaerobie gramnegatīvi koki. Ietver Veillonella ģints oportūnistiskās baktērijas.
Beržeja noteicēja 9. grupa. Riketija un hlamīdijas. Trīs dzimtas - Rickettsiaceae, Bartonellaceae un Chlamydiaceae, no kurām katra satur cilvēkiem patogēnas sugas.
Burgey rokasgrāmatas 10. un 11. grupā ietilpst anoksi- un skābekļa fototrofiskās baktērijas, kas nav patogēnas cilvēkiem.
Burgey determinanta 12. grupa. Aerobās hemolitotrofās baktērijas un radniecīgi organismi. Tas apvieno sēru-dzelzi un mangānu oksidējošās un nitrificējošās baktērijas, kas nerada kaitējumu cilvēkiem.
Burgey rokasgrāmatas 13. un 14. grupā ietilpst pumpurīgās un/vai izaugušās baktērijas un apvalku veidojošās baktērijas. Pārstāv brīvi dzīvojošas sugas, kas nav patogēnas cilvēkiem;
Burgey's guide 15. un 16. grupa apvieno slīdošās baktērijas, kas neveido augļķermeņus un veido tos. Grupās nav iekļautas cilvēkiem patogēnas sugas.
Burgey noteicēja 17. grupa. Grampozitīvi koki. Ietver oportūnistiskās Enterococcus Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus ģints sugas.
Burgey noteicēja 18. grupa. Sporas veidojoši grampozitīvi nūjiņas un koki. Ietver Clostridium un Bacillus ģints patogēnos, nosacīti patogēnos stieņus.
Burgey noteicēja 19. grupa. Sporas veidojošas regulāras formas grampozitīvas nūjiņas. Tostarp oportūnistiskās Erysipelothrix un Listeria ģints sugas.
Burgey noteicēja 20. grupa. Neregulāras formas sporas veidojoši grampozitīvi stieņi. Grupā ietilpst patogēnās un oportūnistiskās ģints Actinomyces, Corynebacterium Gardnerella, Mobiluncus u.c.
Burgey noteicēja 21. grupa. Mikobaktērijas. Ietver vienīgo Mycobacterium ģints, kas apvieno patogēnās un oportūnistiskās sugas.
Grupas 22-29. Aktinomicīti. No daudzām sugām tikai Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi un Terrabacter ģints nekardioformie aktinomicīti (22. grupa) spēj izraisīt bojājumus cilvēkiem.
Burgey determinanta 30. grupa. Mikoplazmas. Acholeplasma, Mycoplasma un Ureaplasma ģints iekļautās sugas ir patogēnas cilvēkiem.
Pārējās Beržeja noteicēju grupas - metanogēnās baktērijas (31), sulfātus reducējošās baktērijas (32 ārkārtīgi halofīlas aerobās arhebaktērijas (33), arhebaktērijas bez šūnu sieniņas (34), ekstrēmi termofīlās un hipertermofīlās, sēru metabolizējošās baktērijas (35) - nav. satur cilvēkiem patogēnas sugas.
Baktērijas ir senākā organismu grupa, kas šobrīd pastāv uz Zemes. Pirmās baktērijas, iespējams, parādījās pirms vairāk nekā 3,5 miljardiem gadu un gandrīz miljardu gadu bija vienīgās dzīvās būtnes uz mūsu planētas. Tā kā šie bija pirmie savvaļas dzīvnieku pārstāvji, viņu ķermenim bija primitīva struktūra.
Laika gaitā to struktūra kļuva sarežģītāka, taču arī mūsdienās baktērijas tiek uzskatītas par primitīvākajiem vienšūnu organismiem. Interesanti, ka dažas baktērijas joprojām saglabā savu seno senču primitīvās iezīmes. To novēro baktērijās, kas dzīvo karstos sēravotos un bezskābekļa dūņās rezervuāru apakšā.
Lielākā daļa baktēriju ir bezkrāsainas. Tikai daži ir krāsoti purpursarkanā vai zaļā krāsā. Bet daudzu baktēriju kolonijām ir spilgta krāsa, kas ir saistīta ar krāsainas vielas izdalīšanos vidē vai šūnu pigmentāciju.
Baktēriju pasaules atklājējs bija 17. gadsimta holandiešu dabaszinātnieks Entonijs Lēvenhuks, kurš pirmais radīja perfektu palielināmo stikla mikroskopu, kas objektus palielina 160-270 reizes.
Baktērijas tiek klasificētas kā prokariotes un tiek sadalītas atsevišķā valstībā - Baktērijas.
ķermeņa forma
Baktērijas ir daudz un dažādi organismi. Tie atšķiras pēc formas.
| baktērijas nosaukums | Baktēriju forma | Baktēriju attēls |
| cocci | sfērisks | |
| Bacillus |  | stieņa formas |
| Vibrio | izliekts komats | |
| Spirillum |  | Spirāle |
| streptokoki |  | Cocci ķēde |
| Stafilokoki |  | Koku kopas |
| diplokoki | Divas apaļas baktērijas, kas ievietotas vienā gļotainā kapsulā |
Pārvietošanās veidi
Starp baktērijām ir mobilās un nekustīgās formas. Kustīgie pārvietojas ar viļņveidīgām kontrakcijām vai ar flagellu (savīti spirālveida pavedieni) palīdzību, kas sastāv no īpaša flagellīna proteīna. Var būt viena vai vairākas flagellas. Dažās baktērijās tās atrodas vienā šūnas galā, citās - divās vai pa visu virsmu.

Taču kustība ir raksturīga arī daudzām citām baktērijām, kurām nav flagellas. Tātad baktērijas, kas no ārpuses pārklātas ar gļotām, spēj slīdēt.
Dažām ūdens un augsnes baktērijām bez flagellas citoplazmā ir gāzes vakuoli. Šūnā var būt 40-60 vakuolu. Katrs no tiem ir piepildīts ar gāzi (domājams, slāpekli). Regulējot gāzes daudzumu vakuolos, ūdens baktērijas var iegrimt ūdens stabā vai pacelties uz tās virsmu, savukārt augsnes baktērijas var pārvietoties augsnes kapilāros.
Dzīvotne
Organizācijas vienkāršības un nepretenciozitātes dēļ baktērijas dabā ir plaši izplatītas. Baktērijas atrodas visur: pat vistīrākā avota ūdens pilē, augsnes graudos, gaisā, akmeņos, polārajos sniegos, tuksneša smiltīs, okeāna dibenā, eļļā, kas iegūta no liela dziļuma, un pat karstā ūdenī. avota ūdens ar temperatūru aptuveni 80ºС. Viņi dzīvo uz augiem, augļiem, dažādiem dzīvniekiem un cilvēkiem zarnās, mutē, ekstremitātēs un uz ķermeņa virsmas.
Baktērijas ir mazākās un daudzskaitlīgākās dzīvās būtnes. Mazā izmēra dēļ tie viegli iekļūst jebkurās plaisās, spraugās, porās. Ļoti izturīgs un pielāgots dažādiem eksistences apstākļiem. Viņi panes žāvēšanu, lielu aukstumu, karsēšanu līdz 90ºС, nezaudējot dzīvotspēju.
Uz Zemes praktiski nav vietas, kur baktērijas netiktu atrastas, bet dažādos daudzumos. Baktēriju dzīves apstākļi ir dažādi. Dažiem no tiem ir nepieciešams gaisa skābeklis, citiem tas nav vajadzīgs un spēj dzīvot bezskābekļa vidē.
Gaisā: baktērijas paceļas augšējos atmosfēras slāņos līdz 30 km. un vēl.
Īpaši daudz no tiem augsnē. Viens grams augsnes var saturēt simtiem miljonu baktēriju.
Ūdenī: atklāto rezervuāru virszemes ūdens slāņos. Noderīgās ūdens baktērijas mineralizē organiskās atliekas.
Dzīvos organismos: patogēnās baktērijas nokļūst organismā no ārējās vides, bet tikai labvēlīgos apstākļos izraisa slimības. Simbiotiķi dzīvo gremošanas orgānos, palīdzot sadalīt un asimilēt pārtiku, sintezēt vitamīnus.
Ārējā struktūra
Baktērijas šūna ir ietērpta īpašā blīvā apvalkā - šūnas sieniņā, kas veic aizsargfunkcijas un atbalsta funkcijas, kā arī piešķir baktērijai paliekošu, raksturīgu formu. Baktērijas šūnu siena atgādina augu šūnas apvalku. Tas ir caurlaidīgs: caur to barības vielas brīvi nokļūst šūnā, un vielmaiņas produkti iziet vidē. Baktērijas bieži veido papildu aizsargājošu gļotu slāni, kapsulu virs šūnas sienas. Kapsulas biezums var būt daudzkārt lielāks par pašas šūnas diametru, taču tas var būt ļoti mazs. Kapsula nav obligāta šūnas sastāvdaļa, tā veidojas atkarībā no apstākļiem, kādos nonāk baktērijas. Tas neļauj baktērijām izžūt.
Dažu baktēriju virspusē ir garas karogs (viena, divas vai daudzas) vai īsas plānas bārkstiņas. Ziedu garums var būt daudzkārt lielāks par baktērijas ķermeņa izmēru. Baktērijas pārvietojas ar flagellas un bārkstiņu palīdzību.

Iekšējā struktūra
Baktēriju šūnas iekšpusē ir blīva nekustīga citoplazma. Tam ir slāņaina struktūra, tajā nav vakuolu, tāpēc dažādas olbaltumvielas (enzīmi) un rezerves barības vielas atrodas pašā citoplazmas vielā. Baktēriju šūnām nav kodola. Viņu šūnu centrālajā daļā ir koncentrēta viela, kas satur iedzimtu informāciju. Baktērijas, - nukleīnskābe - DNS. Bet šī viela nav ierāmēta kodolā.
Baktēriju šūnas iekšējā organizācija ir sarežģīta, un tai ir savas īpatnības. Citoplazmu no šūnas sienas atdala citoplazmas membrāna. Citoplazmā izšķir galveno vielu jeb matricu, ribosomas un nelielu skaitu membrānu struktūru, kas veic dažādas funkcijas (mitohondriju analogi, endoplazmatiskais tīkls, Golgi aparāts). Baktēriju šūnu citoplazmā bieži ir dažādas formas un izmēra granulas. Granulas var sastāvēt no savienojumiem, kas kalpo kā enerģijas un oglekļa avots. Tauku pilieni ir atrodami arī baktēriju šūnā.
Šūnas centrālajā daļā kodolviela DNS ir lokalizēta, nevis ar membrānu atdalīta no citoplazmas. Tas ir kodola analogs - nukleoīds. Nukleoīdam nav membrānas, kodola un hromosomu kopas.
Uztura metodes
Baktērijām ir dažādi barošanas veidi. Starp tiem ir autotrofi un heterotrofi. Autotrofi ir organismi, kas var patstāvīgi veidot organiskas vielas savai uzturam.
Slāpeklis augiem ir nepieciešams, bet tie paši nevar absorbēt slāpekli no gaisa. Dažas baktērijas apvieno gaisā esošās slāpekļa molekulas ar citām molekulām, kā rezultātā tiek iegūtas augiem pieejamas vielas.
Šīs baktērijas apmetas jauno sakņu šūnās, kā rezultātā uz saknēm veidojas sabiezējumi, ko sauc par mezgliņiem. Šādi mezgliņi veidojas uz pākšaugu dzimtas augu un dažu citu augu saknēm.

Saknes nodrošina baktērijas ar ogļhidrātiem, bet baktērijas dod saknēm slāpekli saturošas vielas, kuras augs var uzņemt. Viņu attiecības ir abpusēji izdevīgas.
Augu saknes izdala daudzas organiskas vielas (cukurus, aminoskābes un citas), ar kurām barojas baktērijas. Tāpēc īpaši daudz baktēriju apmetas augsnes slānī, kas ieskauj saknes. Šīs baktērijas pārvērš mirušās augu atliekas augam pieejamās vielās. Šo augsnes slāni sauc par rizosfēru.
Pastāv vairākas hipotēzes par mezgliņu baktēriju iekļūšanu sakņu audos:
- caur epidermas un garozas audu bojājumiem;
- caur sakņu matiņiem;
- tikai caur jauno šūnu membrānu;
- kompanjonu baktēriju dēļ, kas ražo pektinolītiskos enzīmus;
- B-indoletiķskābes sintēzes stimulēšanas dēļ no triptofāna, kas vienmēr atrodas augu sakņu izdalījumos.
Mezglu baktēriju ievadīšanas process sakņu audos sastāv no divām fāzēm:
- sakņu matiņu infekcija;
- mezglu veidošanās process.
Vairumā gadījumu invāzijas šūna aktīvi vairojas, veido tā sauktos infekcijas pavedienus un jau šādu pavedienu veidā pārvietojas augu audos. Mezglu baktērijas, kas izcēlušās no infekcijas pavediena, turpina vairoties saimniekaudi.

Piepildītas ar strauji vairojošām mezgla baktēriju šūnām, augu šūnas sāk intensīvi dalīties. Jauna mezgliņa savienojums ar pākšaugu sakni tiek veikts, pateicoties asinsvadu-šķiedru saišķiem. Funkcionēšanas periodā mezgliņi parasti ir blīvi. Līdz optimālas aktivitātes izpausmes brīdim mezgliņi iegūst rozā krāsu (legoglobīna pigmenta dēļ). Tikai tās baktērijas, kas satur legoglobīnu, spēj piesaistīt slāpekli.
Mezglu baktērijas rada desmitiem un simtiem kilogramu slāpekļa mēslojuma uz vienu hektāru augsnes.
Vielmaiņa
Baktērijas atšķiras viena no otras metabolismā. Dažiem tas notiek ar skābekļa līdzdalību, citiem - bez tā līdzdalības.
Lielākā daļa baktēriju barojas ar gatavām organiskām vielām. Tikai dažas no tām (zili zaļas jeb zilaļģes) spēj radīt organiskas vielas no neorganiskām. Viņiem bija svarīga loma skābekļa uzkrāšanā Zemes atmosfērā.
Baktērijas absorbē vielas no ārpuses, saplēš to molekulas, no šīm daļām saliek apvalku un papildina to saturu (tā tās aug), un izmet nevajadzīgās molekulas. Baktērijas apvalks un membrāna ļauj tai absorbēt tikai pareizās vielas.

Ja baktērijas apvalks un membrāna būtu pilnībā necaurlaidīgi, šūnā neiekļūtu nekādas vielas. Ja tie būtu caurlaidīgi visām vielām, šūnas saturs sajauktos ar barotni – šķīdumu, kurā dzīvo baktērija. Baktēriju izdzīvošanai ir nepieciešams apvalks, kas laiž cauri nepieciešamās vielas, bet ne tās, kuras nav vajadzīgas.
Baktērija absorbē barības vielas, kas atrodas tās tuvumā. Kas notiek tālāk? Ja tas var pārvietoties patstāvīgi (pārvietojot zizli vai atgrūžot gļotas), tad tas kustas, līdz atrod nepieciešamās vielas.

Ja tas nevar kustēties, tad gaida, kamēr difūzija (vienas vielas molekulu spēja iekļūt citas vielas molekulu biezumā) atnes tai nepieciešamās molekulas.
Baktērijas kopā ar citām mikroorganismu grupām veic milzīgu ķīmisku darbu. Pārveidojot dažādus savienojumus, tie saņem savai dzīvībai nepieciešamo enerģiju un uzturvielas. Vielmaiņas procesi, enerģijas iegūšanas veidi un nepieciešamība pēc materiāliem sava organisma vielu veidošanai baktērijās ir dažādi.
Citas baktērijas apmierina visas oglekļa vajadzības, kas nepieciešamas ķermeņa organisko vielu sintēzei uz neorganisko savienojumu rēķina. Tos sauc par autotrofiem. Autotrofās baktērijas spēj sintezēt organiskās vielas no neorganiskām. Starp tiem izšķir:
Ķīmijsintēze
Starojuma enerģijas izmantošana ir vissvarīgākais, bet ne vienīgais veids, kā radīt organiskās vielas no oglekļa dioksīda un ūdens. Ir zināmas baktērijas, kas kā enerģijas avots šādai sintēzei izmanto nevis saules gaismu, bet gan ķīmisko saišu enerģiju, kas rodas organismu šūnās noteiktu neorganisko savienojumu oksidēšanās laikā - sērūdeņradi, sēru, amonjaku, ūdeņradi, slāpekļskābi, dzelzs savienojumus. dzelzs un mangāns. Viņi izmanto organiskās vielas, kas veidojas, izmantojot šo ķīmisko enerģiju, lai izveidotu sava ķermeņa šūnas. Tāpēc šo procesu sauc par ķīmijsintēzi.
Vissvarīgākā ķīmiski sintētisko mikroorganismu grupa ir nitrificējošās baktērijas. Šīs baktērijas dzīvo augsnē un veic amonjaka, kas veidojas organisko atlieku sabrukšanas laikā, oksidēšanu līdz slāpekļskābei. Pēdējais, reaģējot ar augsnes minerālu savienojumiem, pārvēršas slāpekļskābes sāļos. Šis process notiek divos posmos.
Dzelzs baktērijas pārvērš dzelzi par oksīdu. Izveidotais dzelzs hidroksīds nosēžas un veido tā saukto purva dzelzsrūdu.
Daži mikroorganismi pastāv molekulārā ūdeņraža oksidēšanās dēļ, tādējādi nodrošinot autotrofisku uztura veidu.
Ūdeņraža baktērijām raksturīga iezīme ir spēja pāriet uz heterotrofisku dzīvesveidu, ja tās tiek nodrošinātas ar organiskiem savienojumiem un ja nav ūdeņraža.
Tādējādi ķīmijautotrofi ir tipiski autotrofi, jo tie patstāvīgi sintezē nepieciešamos organiskos savienojumus no neorganiskām vielām un neņem tos gatavus no citiem organismiem, piemēram, heterotrofiem. Ķīmijautotrofās baktērijas atšķiras no fototrofiskajiem augiem ar to pilnīgu neatkarību no gaismas kā enerģijas avota.
baktēriju fotosintēze
Dažas pigmentu saturošas sēra baktērijas (violeta, zaļa), kas satur specifiskus pigmentus - bakteriohlorofilus, spēj absorbēt saules enerģiju, ar kuras palīdzību sērūdeņradis tiek sadalīts to organismos un dod ūdeņraža atomus, lai atjaunotu atbilstošos savienojumus. Šim procesam ir daudz kopīga ar fotosintēzi un tas atšķiras tikai ar to, ka purpursarkanajās un zaļajās baktērijās sērūdeņradis (reizēm karbonskābes) ir ūdeņraža donors, bet zaļajos augos tas ir ūdens. Tajos un citos ūdeņraža sadalīšana un pārvietošana tiek veikta absorbēto saules staru enerģijas dēļ.
Šādu baktēriju fotosintēzi, kas notiek bez skābekļa izdalīšanās, sauc par fotoreducēšanu. Oglekļa dioksīda fotoreducēšana ir saistīta ar ūdeņraža pārnešanu nevis no ūdens, bet no sērūdeņraža:
6CO 2 + 12H 2 S + hv → C6H 12 O 6 + 12S \u003d 6H 2 O
Ķīmosintēzes un baktēriju fotosintēzes bioloģiskā nozīme planētu mērogā ir salīdzinoši neliela. Sēra ciklā dabā nozīmīgu lomu spēlē tikai ķīmiskās sintētiskās baktērijas. Uzsūcas zaļajos augos sērskābes sāļu veidā, sērs tiek atjaunots un kļūst par olbaltumvielu molekulu daļu. Turklāt, kad putrefaktīvās baktērijas iznīcina mirušās augu un dzīvnieku atliekas, sērs izdalās sērūdeņraža veidā, ko sēra baktērijas oksidē līdz brīvam sēram (vai sērskābei), kas augsnē veido augiem pieejamus sulfītus. Ķīmiskās un fotoautotrofās baktērijas ir būtiskas slāpekļa un sēra ciklā.
sporulācija
Sporas veidojas baktēriju šūnas iekšpusē. Sporu veidošanās procesā baktēriju šūnā notiek virkne bioķīmisko procesu. Brīvā ūdens daudzums tajā samazinās, fermentatīvā aktivitāte samazinās. Tas nodrošina sporu izturību pret nelabvēlīgiem vides apstākļiem (augsta temperatūra, augsta sāls koncentrācija, žāvēšana utt.). Sporu veidošanās ir raksturīga tikai nelielai baktēriju grupai.

Sporas nav būtisks baktēriju dzīves cikla posms. Sporulācija sākas tikai ar barības vielu trūkumu vai vielmaiņas produktu uzkrāšanos. Baktērijas sporu veidā ilgstoši var palikt miera stāvoklī. Baktēriju sporas iztur ilgstošu vārīšanu un ļoti ilgu sasalšanu. Iestājoties labvēlīgiem apstākļiem, strīds uzdīgst un kļūst dzīvotspējīgs. Baktēriju sporas ir pielāgojumi izdzīvošanai nelabvēlīgos apstākļos.
pavairošana
Baktērijas vairojas, sadalot vienu šūnu divās daļās. Sasniedzot noteiktu izmēru, baktērija sadalās divās identiskās baktērijās. Tad katrs no tiem sāk baroties, aug, dalās utt.

Pēc šūnas pagarināšanas pakāpeniski veidojas šķērseniskā starpsiena, un tad meitas šūnas atšķiras; daudzās baktērijās noteiktos apstākļos šūnas pēc dalīšanās paliek savienotas raksturīgās grupās. Šajā gadījumā atkarībā no dalīšanas plaknes virziena un dalījumu skaita rodas dažādas formas. Reprodukcija ar pumpuru veidošanos notiek baktērijās kā izņēmums.
Labvēlīgos apstākļos šūnu dalīšanās daudzās baktērijās notiek ik pēc 20-30 minūtēm. Ar tik strauju vairošanos vienas baktērijas pēcnācēji 5 dienās spēj izveidot masu, kas spēj piepildīt visas jūras un okeānus. Vienkāršs aprēķins parāda, ka dienā var izveidoties 72 paaudzes (720 000 000 000 000 000 000 šūnas). Ja pārrēķina svarā - 4720 tonnas. Tomēr dabā tas nenotiek, jo lielākā daļa baktēriju ātri iet bojā saules gaismas, izžūšanas, barības trūkuma, uzkarsēšanas līdz 65-100ºС ietekmē, sugu cīņas rezultātā utt.
Baktērija (1), uzsūkusi pietiekami daudz pārtikas, palielinās (2) un sāk gatavoties reprodukcijai (šūnu dalīšanās). Tās DNS (baktērijās DNS molekula ir noslēgta gredzenā) dubultojas (baktērija rada šīs molekulas kopiju). Šķiet, ka abas DNS molekulas (3.4.) ir piestiprinātas pie baktēriju sienas, un, ja tās ir izstieptas, baktērijas novirzās uz sāniem (5.6.). Pirmkārt, sadalās nukleotīds, pēc tam citoplazma.
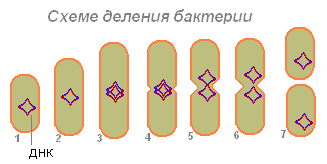
Pēc divu DNS molekulu novirzīšanās uz baktērijām parādās sašaurināšanās, kas pakāpeniski sadala baktērijas ķermeni divās daļās, no kurām katra satur DNS molekulu (7).
Tā notiek (siena bacilis), divas baktērijas salīp kopā, un starp tām veidojas tilts (1,2).

DNS tiek transportēta no vienas baktērijas uz otru, izmantojot džemperi (3). Nokļūstot vienā baktērijā, DNS molekulas savijas, dažviet salīp kopā (4), pēc tam apmainās ar sekcijām (5).
Baktēriju loma dabā
Aprite
Baktērijas ir vissvarīgākā saikne vispārējā vielu apritē dabā. Augi veido sarežģītas organiskas vielas no oglekļa dioksīda, ūdens un augsnes minerālsāļiem. Šīs vielas atgriežas augsnē ar mirušām sēnītēm, augiem un dzīvnieku līķiem. Baktērijas sadala sarežģītas vielas vienkāršās, kuras atkārtoti izmanto augi.
Baktērijas iznīcina mirušo augu un dzīvnieku līķu sarežģītās organiskās vielas, dzīvo organismu ekskrēcijas un dažādus atkritumus. Barojot ar šīm organiskajām vielām, saprofītiskās pūšanas baktērijas tās pārvērš humusā. Tādi ir mūsu planētas sakārtotāji. Tādējādi baktērijas aktīvi iesaistās vielu apritē dabā.
augsnes veidošanās
Tā kā baktērijas ir izplatītas gandrīz visur un ir sastopamas milzīgā daudzumā, tās lielā mērā nosaka dažādus dabā notiekošos procesus. Rudenī kokiem un krūmiem krīt lapas, nokalst virszemes zāles dzinumi, nobirst vecie zari, ik pa laikam nobirst veco koku stumbri. Tas viss pamazām pārvēršas humusā. 1 cm3. Meža augsnes virskārtā ir simtiem miljonu vairāku sugu saprofītu augsnes baktēriju. Šīs baktērijas pārvērš humusu dažādās minerālvielās, kuras no augsnes var absorbēt augu saknes.
Dažas augsnes baktērijas spēj absorbēt slāpekli no gaisa, izmantojot to dzīvības procesos. Šīs slāpekli fiksējošās baktērijas dzīvo pašas vai apmetas pākšaugu saknēs. Iekļūstot pākšaugu saknēs, šīs baktērijas izraisa sakņu šūnu augšanu un mezgliņu veidošanos uz tām.
Šīs baktērijas izdala slāpekļa savienojumus, ko izmanto augi. Baktērijas iegūst ogļhidrātus un minerālsāļus no augiem. Tādējādi starp pākšaugu un mezglu baktērijām pastāv cieša saistība, kas ir noderīga gan vienam, gan otram organismam. Šo parādību sauc par simbiozi.
Pateicoties simbiozei ar mezgliņu baktērijām, pākšaugi bagātina augsni ar slāpekli, palīdzot palielināt ražu.
Izplatība dabā
Mikroorganismi ir visuresoši. Vienīgie izņēmumi ir aktīvo vulkānu krāteri un nelielas teritorijas detonēto atombumbu epicentros. Ne Antarktikas zemās temperatūras, ne geizeru vārošās strūklas, ne piesātinātie sāls šķīdumi sāls baseinos, ne kalnu virsotņu spēcīgā insolācija, ne kodolreaktoru skarbais starojums netraucē mikrofloras pastāvēšanu un attīstību. Visas dzīvās būtnes pastāvīgi mijiedarbojas ar mikroorganismiem, bieži vien ir ne tikai to glabātāji, bet arī izplatītāji. Mikroorganismi ir mūsu planētas pamatiedzīvotāji, aktīvi attīstot neticamākos dabiskos substrātus.
Augsnes mikroflora
Baktēriju skaits augsnē ir ārkārtīgi liels - simtiem miljonu un miljardu īpatņu 1 gramā. Augsnē tie ir daudz vairāk nekā ūdenī un gaisā. Kopējais baktēriju skaits augsnēs ir atšķirīgs. Baktēriju skaits ir atkarīgs no augsnes veida, to stāvokļa, slāņu dziļuma.
Uz augsnes daļiņu virsmas mikroorganismi atrodas nelielās mikrokolonijās (katrā pa 20-100 šūnām). Bieži tie veidojas organisko vielu recekļu biezumos, uz dzīvām un mirstošām augu saknēm, tievos kapilāros un iekšā kunkuļos.
Augsnes mikroflora ir ļoti daudzveidīga. Šeit sastopamas dažādas baktēriju fizioloģiskās grupas: pūšanas, nitrificējošās, slāpekli fiksējošās, sēra baktērijas uc starp tām ir aerobās un anaerobās, sporu un nesporu formas. Mikroflora ir viens no augsnes veidošanās faktoriem.
Mikroorganismu attīstības zona augsnē ir zona, kas atrodas blakus dzīvo augu saknēm. To sauc par rizosfēru, un tajā esošo mikroorganismu kopumu sauc par rizosfēras mikrofloru.
Rezervuāru mikroflora
Ūdens ir dabiska vide, kurā mikroorganismi vairojas lielā skaitā. Lielākā daļa no tiem nonāk ūdenī no augsnes. Faktors, kas nosaka baktēriju skaitu ūdenī, barības vielu klātbūtni tajā. Tīrākie ir artēzisko aku un avotu ūdeņi. Atvērtās ūdenskrātuves un upes ir ļoti bagātas ar baktērijām. Visvairāk baktēriju atrodas ūdens virszemes slāņos, tuvāk krastam. Palielinoties attālumam no krasta un palielinoties dziļumam, baktēriju skaits samazinās.
Tīrā ūdenī ir 100-200 baktēriju uz 1 ml, savukārt piesārņotā ūdenī ir 100-300 tūkstoši un vairāk. Apakšējās dūņās ir daudz baktēriju, it īpaši virsmas slānī, kur baktērijas veido plēvīti. Šajā plēvē ir daudz sēra un dzelzs baktēriju, kas oksidē sērūdeņradi līdz sērskābei un tādējādi novērš zivju bojāeju. Dūņos ir vairāk sporu nesošo formu, savukārt ūdenī dominē sporu nesošās formas.
Pēc sugu sastāva ūdens mikroflora ir līdzīga augsnes mikroflorai, taču sastopamas arī specifiskas formas. Iznīcinot dažādus ūdenī iekritušos atkritumus, mikroorganismi pamazām veic tā saukto bioloģisko ūdens attīrīšanu.
Gaisa mikroflora
Gaisa mikroflora ir mazāka nekā augsnes un ūdens mikroflora. Baktērijas paceļas gaisā kopā ar putekļiem, var tur kādu laiku palikt, bet pēc tam nosēsties uz zemes virsmas un iet bojā no uztura trūkuma vai ultravioleto staru ietekmē. Mikroorganismu skaits gaisā ir atkarīgs no ģeogrāfiskā apgabala, atrašanās vietas, gadalaika, putekļu piesārņojuma utt. Katrs putekļu plankums ir mikroorganismu nesējs. Lielākā daļa baktēriju gaisā atrodas virs rūpniecības uzņēmumiem. Laukos gaiss ir tīrāks. Tīrākais gaiss ir virs mežiem, kalniem, sniegotām vietām. Gaisa augšējos slāņos ir mazāk baktēriju. Gaisa mikroflorā ir daudz pigmentētu un sporu nesēju baktēriju, kas ir izturīgākas par citām pret ultravioletajiem stariem.
Cilvēka ķermeņa mikroflora
Cilvēka ķermenis, pat pilnīgi vesels, vienmēr ir mikrofloras nesējs. Cilvēka ķermenim saskaroties ar gaisu un augsni, uz apģērba un ādas nogulsnējas dažādi mikroorganismi, tostarp patogēni (stingumkrampju baciļi, gāzes gangrēna u.c.). Visbiežāk tiek piesārņotas atklātās cilvēka ķermeņa daļas. Uz rokām atrodami E. coli, stafilokoki. Mutes dobumā ir vairāk nekā 100 veidu mikrobu. Mute ar savu temperatūru, mitrumu, barības vielu atliekām ir lieliska vide mikroorganismu attīstībai.
Kuņģī notiek skāba reakcija, tāpēc lielākā daļa tajā esošo mikroorganismu mirst. Sākot no tievās zarnas, reakcija kļūst sārmaina, t.i. labvēlīgs mikrobiem. Mikroflora resnajā zarnā ir ļoti daudzveidīga. Katrs pieaugušais katru dienu ar ekskrementiem izdala aptuveni 18 miljardus baktēriju, t.i. vairāk cilvēku nekā cilvēku uz zemeslodes.
Iekšējie orgāni, kas nav saistīti ar ārējo vidi (smadzenes, sirds, aknas, urīnpūslis utt.), parasti ir brīvi no mikrobiem. Mikrobi šajos orgānos iekļūst tikai slimības laikā.
Baktērijas riteņbraukšanā
Mikroorganismiem kopumā un jo īpaši baktērijām ir liela nozīme bioloģiski svarīgajos vielu ciklos uz Zemes, veicot ķīmiskas transformācijas, kas ir pilnīgi nepieejamas ne augiem, ne dzīvniekiem. Dažādus elementu cikla posmus veic dažāda veida organismi. Katras atsevišķas organismu grupas pastāvēšana ir atkarīga no elementu ķīmiskās transformācijas, ko veic citas grupas.
slāpekļa cikls
Slāpekļa savienojumu cikliskajai transformācijai ir vissvarīgākā loma, nodrošinot nepieciešamo slāpekļa formu dažādiem biosfēras organismiem uztura vajadzību ziņā. Vairāk nekā 90% no kopējā slāpekļa fiksācijas notiek dažu baktēriju metaboliskās aktivitātes dēļ.
Oglekļa cikls
Organiskā oglekļa bioloģiskai pārvēršanai oglekļa dioksīdā, ko pavada molekulārā skābekļa samazināšanās, ir nepieciešama dažādu mikroorganismu kopīga vielmaiņas darbība. Daudzas aerobās baktērijas veic pilnīgu organisko vielu oksidēšanu. Aerobos apstākļos organiskie savienojumi sākotnēji tiek sadalīti fermentācijas ceļā, un organiskās fermentācijas galaprodukti tiek tālāk oksidēti anaerobās elpošanas ceļā, ja ir neorganiskie ūdeņraža akceptori (nitrāti, sulfāti vai CO2).
Sēra cikls
Dzīviem organismiem sērs ir pieejams galvenokārt šķīstošu sulfātu vai reducētu organisko sēra savienojumu veidā.
Dzelzs cikls
Dažās saldūdens rezervuāros ir liela reducēto dzelzs sāļu koncentrācija. Šādās vietās veidojas specifiska baktēriju mikroflora - dzelzs baktērijas, kas oksidē reducēto dzelzi. Tie piedalās purva dzelzsrūdu un ar dzelzs sāļiem bagātu ūdens avotu veidošanā.
Baktērijas ir senākie organismi, kas arhejā parādījās apmēram pirms 3,5 miljardiem gadu. Apmēram 2,5 miljardus gadu viņi dominēja uz Zemes, veidojot biosfēru, un piedalījās skābekļa atmosfēras veidošanā.
Baktērijas ir vieni no visvienkāršāk sakārtotajiem dzīviem organismiem (izņemot vīrusus). Tiek uzskatīts, ka tie ir pirmie organismi, kas parādījās uz Zemes.
baktērijas- Tie ir vienšūnas, bez hlorofila organismi, kas vairojas veģetatīvi daloties, retāk šņorējot, dažkārt veidojot intracelulāras sporas.
Baktēriju lielumu mēra mikronos un, ar retiem izņēmumiem, svārstās no 0,06-0,3 līdz 3-5 μ. Ūdens pile var viegli saturēt vairākus simtus miljonu baktēriju.
Baktēriju šūnas forma ir diezgan viendabīga. Ir zināmas trīs galvenās baktēriju formas: apaļas, stieņa formas un savītas ar daudzām un neuzkrītošām pārejām starp tām. Antons De Bari tās tēlaini salīdzināja ar biljarda bumbu, zīmuli un korķviļķi.
Baktērijas, kurām ir apaļa forma, sauc par koku. Tie atšķiras pēc izmēra un relatīvā stāvokļa. Kokus, kas savienoti pa pāriem, sauc par diplokokiem, bet savienotus kaklarotas veidā sauc par streptokokiem. Daloties, pārmaiņus divos savstarpēji perpendikulāros virzienos, veidojas tetrakoki. Ja dalījums ir pareizs un atkārtojas trīs savstarpēji perpendikulāros virzienos, tad šūnu savienojumi veidojas iepakojumu veidā – tās ir tā saucamās sardīnes. Sadaloties dažādos virzienos bez īpašas regularitātes, koki veido nejaušas kopas, kas atgādina vīnogu suku. Tos sauc par stafilokokiem.
Stieņveida baktērijas pēc izskata ir nedaudz daudzveidīgākas. Tās var būt ar nošķeltiem vai noapaļotiem galiem, cilindriskas, mucveida vai citrona formas un it kā ar sašaurinājumu vidū, bieži vien elipsoidālas, atšķiras tikai ar platumu un garumu. Spieķi var būt taisni, izliekti, atsevišķi, savienoti pa pāriem vai ķēdē, īsi vai stipri iegareni. Stieņveida baktērijas, kuru garums ir divreiz vai vairāk par platumu, sauc par baciļiem; ja starpība starp garumu un platumu ir maza, tad tās sauc par baktērijām.
Cirtainas formas baktērijas atšķiras ne tikai pēc garuma un biezuma, bet arī pēc cirtas skaita un rakstura. Nedaudz izliektas baktērijas (čokurošanās nepārsniedz 1/4 spirāles apgriezienu) sauc par vibrioniem, baktērijas, kurām ir viena vai vairākas lielas regulāras cirtas, sauc par spirilumu. Garas un plānas izliektas formas baktērijas ar daudzām mazām cirtām, dažreiz ar lieliem visa pavediena izliekumiem, sauc par spirohetām.
Baktēriju struktūra
Savas organizācijas vienkāršības un nenozīmīgā izmēra dēļ baktērijas pieder pie visvienkāršākajām būtnēm un stāv uz zemākajām dzīvības pakāpieniem. Neskatoties uz milzīgajiem zinātnes un tehnikas sasniegumiem, vēl nav atrisināti visi jautājumi par baktēriju struktūru.
Baktēriju ķermenis sastāv no ētiska satura čaumalas un protoplazmas, kas piesūcināta ar šūnu sulu. Baktēriju apvalks ir plāns, bezkrāsains, tā struktūra mikroskopā nav atšķirama. Lai to redzētu, viņi izmanto mākslīgas apstrādes metodes. Apvalks ir šūnas ārējās formas pamatā un, šķiet, ir labi pazīstama aizsardzība pret nelabvēlīgiem apstākļiem. Brīvi aptverot šūnas saturu, pateicoties tās elastībai, tā ļauj brīvi pārvietoties baktērijām, ko bieži pavada enerģiskas visa ķermeņa kustības.
Korpusa ārējie slāņi, uzsūcot ūdeni, bieži uzbriest un veido želatīniski lipīgu masu, sasniedzot pamanāmu izmēru. Tā kā ārējie slāņi kļūst gļotaini, apvalks tiek nepārtraukti atjaunots uz protoplazmas rēķina. Atdzesētu līmes apvalku sauc par kapsulu. Gļotu un kapsulu veidošanās intensitāte ir atkarīga no uztura īpašībām un dažkārt var būt ļoti nozīmīga. Gļotādas kapsula veidojas ne tikai pie katras šūnas atsevišķi, bet arī daudzās šūnās, kas savienotas vienā kolonijā un it kā ietvertas kopējā kapsulā. Šīs gļotainās baktēriju kolonijas sauc zoogles. Kapsulu veidošanās nav raksturīga visiem baktēriju veidiem.
baktēriju kustība
Spēja veikt spontānu kustību ir raksturīga tikai dažām baktēriju grupām. Baktērijas pārvietojas ar karogiem vai skropstiņiem. Flagella izskatās kā vairāk vai mazāk gari pavedieni. Tie ir ļoti delikāti, plāni, viegli nolūst un nav redzami mikroskopā bez īpašas krāsošanas. To diametrs nepārsniedz 1/20 no baktēriju ķermeņa diametra.
Mobilās baktēriju formas iedala šādās grupās:
- vienveidīgs - ir tikai viens polārais karogs,
- lophotrichous - vienā šūnas galā ir karogs saišķis,
- peritrichous flagellas atrodas pa visu ķermeņa virsmu.
Ziedu atrašanās vieta uz baktērijas ķermeņa nosaka tās kustības raksturu - taisnu vai nejaušu. Baktēriju kustīgums ir atkarīgs no vairākiem apstākļiem: temperatūras, barotnes sastāva, to dzīvībai svarīgās aktivitātes produktiem utt. Kustīgās baktēriju formas nav aprīkotas ar flagellas visos to attīstības posmos un ne visos augšanas apstākļos.
sporulācija
Daudzu baktēriju organismā noteiktos to attīstības periodos parādās apaļi vai elipsoidāli veidojumi - balsti. Viņi parasti pabeidz baktēriju attīstības ciklu. Sporu izmērs, salīdzinot ar to šūnu lielumu, kas tās radīja, var ievērojami atšķirties.
Balsti neveidojas visu veidu baktērijās. Tos ieskauj labi izolēts apvalks, gandrīz ūdens necaurlaidīgs, un tie ir visstabilākie veidojumi visā dzīvajā pasaulē. Tādējādi tie bieži iztur vārīšanu vairākas stundas un ilgstošu sausā tvaika iedarbību temperatūrā no 120 līdz 140 °. Daudzu baciļu sporas saglabā savu dzīvotspēju pēc ilgstošas uzturēšanās -190 ° temperatūrā un pat šķidrā ūdeņraža temperatūrā (-253 °). Tie ir arī izturīgi pret ķīmisko vielu iedarbību - indēm. Tas viss apgrūtina cīņu pret patogēnām baktēriju sporu sugām.
Nobriedusi spora var palikt dzīvotspējīga gadu desmitiem. Parasti sporu dīgšana notiek pēc noteikta miera perioda un ir saistīta ar ārējo apstākļu ietekmi. Viss sporulācijas process notiek vienas dienas laikā vai mazāk. Pēc sporu nobriešanas šūna, kas to radījusi, pakāpeniski izmirst un nobriedusi spora izdalās. Dīgšanas laikā tas uzbriest, kļūst bagātāks ar ūdeni, un no tā iznāk stāds, ietērpts plānā čaulā.
Baktēriju pavairošana
Sasniedzot brieduma stāvokli un augšanas robežu, baktērijas sāk vairoties, vienkārši daloties. Dalīšanās laikā baktērijas ķermeņa vidusdaļā parādās starpsiena, kas pēc tam sadalās un atdala divas jaunas šūnas. Starpsienu secīgais izvietojums dalīšanas laikā dažādām baktērijām ir atšķirīgs. Stieņveida formās tas atrodas perpendikulāri garajai asij, sfēriskās formās starpsienas var atrasties vienā, divās vai trīs savstarpēji perpendikulārās plaknēs, kas ir iemesls tādu formu kā streptokoku, tetrakoku un sarkīnu veidošanās. .
Baktēriju vairošanās ātrums ir atkarīgs no vairākiem apstākļiem un var būt ļoti atšķirīgs. Jo labvēlīgāki ārējie apstākļi baktēriju pastāvēšanai, jo ātrāk notiek to sadalīšanās. Normālos apstākļos baktēriju skaits dubultojas aptuveni ik pēc pusstundas. Ja tas vienmēr notiktu netraucēti, tad baktēriju skaits no vienas šūnas sasniegtu kolosālus apmērus. Pēc mikrobiologa Kohna domām, viena baciļa pēcnācēji piecās dienās varētu piepildīt visas jūras un okeānus. Tomēr tas nekad nav noticis un nekad nenotiks. Baktēriju dzīves ciklu ierobežo noteikti ārēji apstākļi, pēc kuriem vairošanās palēninās vai apstājas pavisam. Uztura trūkums, kaitīgi vielmaiņas produkti, dažādu sugu konkurence utt., kaitīgi ietekmē baktērijas. Nelabvēlīgos apstākļos tie masveidā mirst.
Baktēriju klasifikācija
Baktēriju pozīcija dzīvo būtņu sistēmā vēl nav precīzi noteikta. Ir vispāratzīts, ka baktērijas ir daļa no augu pasaules, un sēnes un aļģes ir tām vistuvākie saistītie organismi. Baktēriju morfoloģiskās pazīmes vairumā gadījumu aprobežojas ar dažām formām: sfēriski, stieņi, spirāles. To ārējās organizācijas neparastā vienkāršība un elementārais raksturs apgrūtina to klasificēšanu. Baktērijas sugu noteikt, pamatojoties tikai uz morfoloģiskām pazīmēm, nav iespējams. Zinātniskās sistemātikas pamatā ir morfoloģija un attīstības vēsture, taču, lai klasificētu baktērijas, ir jāzina ne tikai morfoloģija, bet arī to fizioloģiskās un bioķīmiskās īpašības. Šajā sakarā tiek noteikts: baktēriju attiecība pret skābekli, temperatūras apstākļi, pigmenta veidošanās, želatīna sašķidrināšana, skābju un gāzu veidošanās uz cukuriem, piena izmaiņas ar baktēriju augšanu tajā, indola, sērūdeņraža, amonjaka veidošanās, nitrātu reducēšanās par nitrītiem vai brīvu slāpekli. Tomēr ar to ne vienmēr pietiek, lai noteiktu baktēriju veidu.
Ir dažādas baktēriju klasifikācijas sistēmas, taču tās visas ir nosacītas un daudz vairāk vai mazāk no dabiskās klasifikācijas. Šo sistēmu vai vismaz vienas no tām izskatīšana šajā gadījumā nav nepieciešama, pat ja to piemēro fitopatogēnām baktērijām. Jāteic vien, ka šobrīd gandrīz visas fitopatogēnās baktērijas ir apvienotas Pseudomonas, Xanthomonas, Bacterium un Erwinia ģintīs.
Nesen M. V. Gorlenko (1961) ierosināja šādu Eubacteriales klases fitopatogēno baktēriju klasifikācijas sistēmu:
es Mycobacteriaceae dzimta(Chester, 1901) - nekustīgas baktērijas (bez flagellas):
- 1. ģints - Gorynebacterium (Leman and Neumann, 1896) - (grampozitīvas baktērijas;
- 2. ģints Aplanobacterium (Smith, 1905, Geshich, 1956) - gramnegatīvas baktērijas.
II. Pseudomonadaceae dzimta(Wilson et al., 1917) - baktērijas ar flagellas (polāras):
- 1. ģints - Pseudomonas (Migula, 1900) - nekrāsotas un fluorescējošas baktērijas;
- 2. ģints - Xanthomonas (Dawson, 1839) - baktērijas ar krāsainām kolonijām.
III. Baktēriju dzimta(Kon, 1872) - kustīgas baktērijas ar peritrichous flagellas, kas neveido balstus:
- 1. ģints - Baktērija (Ehrenberger, 1828) - nekrāsotas formas, kas neveido pektināzes un protopektināzes;
- 2. ģints - Pectobacterium (Waldy, 1945) - nekrāsotas formas, kas veido pektināzi un protopektināzi;
- 3. ģints - Chromobacterium (Bergonzini, 1881) - krāsainas formas.
IV. Bacillaceae dzimta(Fischer, 1895) - kustīgas baktērijas, sporas veidojošas nūjiņas:
- 1. ģints - Bacillus (Kon, 1832) - šūnas sporu veidošanās laikā neuzbriest vai nedaudz uzbriest;
- 2. ģints - Clostridium (Praznovsky, 1880) - sporu veidošanās laikā šūnas uzbriest.
Iepriekš minētajā sistēmā līdz šim vispārpieņemtā Erwinia ģints ir izlaista. No tā tiek izdalīta īpaša Pectobacterium ģints, kurā ietilpst baktērijas ar peritrichous flagellas un pektolītisko aktivitāti. Tās fitopatogēnās baktērijas, kurām šādas spējas nepiemīt, tiek iedalītas baktēriju ģintī. Šī sistēma, pati par sevi racionāla, ir jauna un ikdienā vēl nav ienākusi, tāpēc šajā darbā pieturamies pie klasifikācijas, kurā sava vieta ierādīta Erviniju ģints. Šis fitopatogēno baktēriju sugas nosaukums tiek plaši izmantots specializētajā literatūrā gan mūsu valstī, gan ārvalstīs.
Baktēriju veida noteikšana, neizmantojot mākslīgās barotnes, nav iespējama. Šajā sakarā var atzīmēt, ka, kultivējot baktērijas, tās veido ļoti raksturīgas kolonijas. Šajā gadījumā, lai spriestu par baktēriju sugām, var izmantot vienu izskatu.
Ja atrodat kļūdu, lūdzu, iezīmējiet teksta daļu un noklikšķiniet uz Ctrl+Enter.
1.5. Mūsdienu baktēriju klasifikācija Mūsdienu baktēriju taksonomijā ir izveidojusies situācija, kas raksturīga arī citu organismu klasifikācijai: ir panākts progress filoģenētiskās klasifikācijas sistēmas izveidē, kas atspoguļo galvenos evolūcijas attīstības virzienus un pārstāvju attiecības. atsevišķi taksoni, bet mākslīgās parādības saglabā savu nozīmi.- tipiskas klasifikācijas, ērtākas mikroorganismu identificēšanai. Pašlaik nav detalizētas prokariotu evolūcijas sistēmas, un, visticamāk, šīs problēmas risinājums nav tuvākās nākotnes jautājums. Prokariotu īpašības morfoloģiskās, fizioloģiskās, bioķīmiskās un ģenētiskās organizācijas jomā norāda uz to, ka tiem nav piemērojami labi izstrādāti principi, kas tiek izmantoti augstāku organismu sistēmas veidošanā. Nekavējoties pie baktēriju taksonomijas problēmas vēsturiskajiem aspektiem, jāatzīmē, ka vispieņemamākā prokariotu filoģenētiskā klasifikācijas sistēma ir sistēma, kuras pamatā ir nukleotīdu secības saskaņošana 16S rRNS. Šī sistēma veidoja pamatu daudzsējumu prokariotu enciklopēdijas 2. izdevumam - Beržeja sistemātiskās bakterioloģijas rokasgrāmatai (Bergey's Manual of Systematic Bacteriology), kuras pirmais sējums tika publicēts 2001. gadā. Šajā darbā visi prokarioti ir sadalīti 26 filoģenētiskas "zaras" (grupas), kuru pamatā ir to 16S-rRNS struktūra; 23 "zarus" pārstāv eubaktērijas, bet trīs - arhebaktērijas. Jāuzsver, ka liela daļa šo filoģenētisko grupu satur prokariotu sugas, kas nav izolētas kā tīrkultūras un tāpēc vēl nav detalizēti pētītas. Šo sugu pārstāvjiem pašlaik ir zināmas tikai 16S rRNS nukleotīdu sekvences. No 23 eibaktēriju grupām divas filoģenētiskās grupas pārstāv grampozitīvas baktērijas, pārējās grupas ir gramnegatīvas. Gramnegatīvās baktērijas sastāv no lielas proteobaktēriju (proteobaktērijas) grupas un 20 citu baktēriju grupām ar šāda veida šūnu sieniņām. 1. tabulā ir sniegts īss proteobaktēriju apraksts, kurām vairuma eikariotu mitohondriji un hloroplasti ir vistuvāk 16S-rRNS sastāvā. 2. Proteobaktērijas ir gramnegatīvu baktēriju grupa, kas ir ļoti neviendabīga morfoloģiskā, fizioloģiskā un bioķīmiskā ziņā. Šīs grupas pārstāvjus raksturo visa veida enerģijas vielmaiņa un uzturs. Lielākajai daļai proteobaktēriju sugu šūnas ir stieņveida, sfēriskas vai vibrioīdas formas, vairojas galvenokārt binārās dalīšanās ceļā, bet dažām sugām raksturīga pumpuru veidošanās un augļķermeņu veidošanās sarežģītā šūnu ciklā. Šajā grupā ir gan kustīgas flagellas, gan nekustīgas baktērijas. Attiecībā uz molekulāro skābekli proteobaktērijas tiek klasificētas kā obligāti aerobi, obligāti un fakultatīvi anaerobi. Proteobaktēriju grupa ir sadalīta piecās apakšgrupās, pamatojoties uz atšķirībām 16S rRNS: alfa, beta, gamma, delta un epsilons. Papildus proteobaktērijām gramnegatīvās eubaktērijas ietver šādas galvenās eubaktēriju grupas: ūdeņraža termofīli, zaļās pavedienveida baktērijas, zaļās sēra baktērijas, cianobaktērijas, spirohetas, citofāgi, bakterioīdi, hlamīdijas, planktomicīti, deinokoki, florobaktērijas, termofleksijas, florobaktērijas, uc Filoģenētiskās grupas grampozitīvās baktērijas - Actinobacteria un Firmicutes. Actinobacteria grupu ("actinomycete filiāle") pārstāv šādas baktēriju ģints ar augstu GC pāru saturu DNS: Geodermatophilus, Frankia, Streptomyces, Arthrobacter, Micrococcus, Actinomyces, Bifidobacterium, Propionibacterium, Actinoplanes, Nocardia, Rhodococcus , mikobaktērijas. Firmicutes grupa (“klostrīdu atzars” - galvenokārt grampozitīvas baktērijas ar zemu GC pāru saturu DNS) sastāv no šādām ģintīm: Clostridium, Lactococcus, Pediococcus, Streptococcus, Enterococcus, Leuconostoc, Listeria, Caryococcus, Staphylococcus, Sarcinacoccus. , Sporosarcina, Bacillus, Desulfotomaculum, Heliobacterium, Mycoplasma, Ureaplasma uc Arhebaktērijas iedala trīs filoģenētiskās grupās: Crenarchaeota, Euryarchaeota un Korarchaeota. Crenarchaeota grupa sastāv no ārkārtīgi termofīlām baktērijām, kuru lielākā daļa pārstāvju metabolizē sēru, daži samazina dzelzs un molibdēna jonus. Euryarchaeota grupā ietilpst obligātas anaerobās metanogēnās arhebaktērijas, kā arī ekstrēmi termofīli un halofīli. Korarchaeota grupu veido arhebaktērijas, kas dzīvo karstos sēravotos. Līdz šim neviens no šīs grupas pārstāvjiem (kuriem ir līdzīga 16S rRNS) nav izolēts kā tīrkultūra, tāpēc to fenotipiskās pazīmes nav pietiekami pētītas. Noslēdzot prokariotu filoģenētisko zaru apskatu, jāatzīmē, ka piedāvātā filoģenētiskā sistēma, kuras pamatā ir tikai viena ribosomu RNS gēna nukleotīdu secību izpēte, nav nekas vairāk kā viena no tehniski ērtajām un izstrādātajām sistēmām daudzu pasūtīšanai. organismus, lai tos identificētu, tāpēc ir loģiski izveidot pareizu baktēriju taksonomiju, tikai ņemot vērā šo pazīmi, nav iespējams. Visvairāk atzītā un lietotā baktēriju fenotipiskā klasifikācija ir sniegta Burgey's Guide to Bacteria devītajā izdevumā. Šajā izdevumā baktērijas ir iedalītas četrās galvenajās kategorijās (nodaļās), pamatojoties uz šūnu robežslāņa struktūru: 1) Gracilicutes (no latīņu cutes - āda, gracilis - plānas) - gramnegatīvas eubaktērijas ar šūnu sieniņām; 2) Firmicutes (no lat. firmus - stiprs) - grampozitīvas eubaktērijas ar šūnu sieniņām; 3) Tenericutes (no latīņu tener - mīksts, maigs) - eubaktērijas, bez šūnu sieniņām; 4) Mendosicutes (no latīņu valodas mendosus - kļūdains) - arhebaktērijas, kuru šūnu sienas atšķiras no citu prokariotu līdzīgām struktūrām. Gracilicutes nodaļā ir dažādas morfoloģijas baktērijas ar gramnegatīvu šūnu sieniņu. Vairošanās notiek galvenokārt ar bināro dalīšanos, dažas baktērijas vairojas ar pumpuru veidošanos. Endospora neveidojas. Lielākā daļa ir kustīgas: ir visa veida baktēriju kustība - ar flagella palīdzību, slīdēšana, locīšana. Šajā nodaļā ietilpst aerobās, anaerobās un fakultatīvās anaerobās baktērijas; fototrofās un ķīmijtrofās baktērijas. Nodaļa ir sadalīta trīs klasēs: Scotobacteria, Oxyphotobacteria, Anoxyphotobacteria. Scotobacteria klasē ietilpst gramnegatīvās baktērijas, kas neizmanto gaismas enerģiju vielmaiņas nolūkiem, bet saņem to tikai redoksreakciju rezultātā. Klases nosaukums cēlies no grieķu valodas sсotos - tumsa. Šī ir lielākā baktēriju klase. Anoksifotobaktēriju klasē ietilpst purpursarkanās baktērijas, zaļās baktērijas un heliobaktērijas, kas veic anoksigēno fotosintēzi (bez molekulārā skābekļa izdalīšanās). Oksifotobaktēriju klasi pārstāv cianobaktērijas un prohlorofīti, kas veic skābekļa fotosintēzi (ar molekulārā skābekļa izdalīšanos). Šis fotosintēzes veids ir līdzīgs fotosintēzei, kas notiek augos. Firmicutes nodaļā ir baktērijas ar grampozitīvu šūnu sieniņu. Šūnām var būt dažādas formas: stieņi, koki, pavedienveida, zarojoši. Daži pārstāvji veido endosporas. Lielākā daļa no tiem ir nekustīgi; kustīgajām formām ir peritrichous flagellas. Nodaļā ietilpst aerobās, anaerobās un fakultatīvās anaerobās baktērijas. Nodaļa sastāv no divām klasēm: Firmibacteria, Tallobacteria. Firmibacteria klase ietver lielu skaitu grampozitīvu baktēriju, kas nesazarojas. Tallobaktēriju klasē ietilpst baktērijas, kuru šūnas spēj "zaroties". Tenericutes nodaļu pārstāv baktērijas, kurām nav šūnu sienas. Šūnu sienas trūkuma dēļ šūnu forma nav nemainīga: vienas sugas tīrkultūrā vienlaikus ir kokosveida, stieņveida, pavedienveida, bumbierveida, diskveida un citas šūnas. Šajā sadaļā iekļauto baktēriju vairošanās notiek ar bināro skaldīšanu, pumpuru veidošanos. Grama traips ir negatīvs. Raksturīga ir mazu koloniju veidošanās, kas kļūst par agaru. Tie var būt saprofītiski, parazitāri vai patogēni. Nodaļu veido viena Mollicutes (mikoplazmas) klase. Mendosicutes nodaļu veido baktērijas ar stingru šūnu sienu, bet kurām trūkst mureīna peptidoglikāna. Lielākā daļa pārstāvju ir stingri anaerobi, no kuriem daudziem ir flagellas. Sugām raksturīga ekoloģiskā un vielmaiņas daudzveidība, spēja dzīvot ekstremālos apstākļos. Nodaļu veido viena klase - Arhebaktērijas. Četru departamentu (galveno kategoriju) ietvaros ir identificētas 35 baktēriju grupas (vai sadaļas), kuras lielākā vai mazākā mērā tiks raksturotas turpmākajās nodaļās. Gracilicutes nodaļai pieder šādas grupas. 1. grupa. Spirohetes. 2. grupa. Aerobās (vai mikroaerofilās), kustīgās, spirālveida (vai vibrioīdās) gramnegatīvās baktērijas. 3. grupa. Nekustīgas vai reti kustīgas gramnegatīvas izliektas baktērijas. 4. grupa. Gramnegatīvi aerobās (vai mikroaerofīlās) nūjiņas un koki. 5. grupa. Izvēles aerobās gramnegatīvās stieņi. 6. grupa. Gramnegatīvi anaerobie taisni, izliekti vai spirālveida stieņi. 7. grupa. Baktērijas, kas veic sēra vai sulfāta disimilācijas reducēšanu. 8. grupa. Anaerobie gramnegatīvi koki. 9. grupa. Riketsija un hlamīdijas. 10. grupa. Anoksigēnās fototrofās baktērijas. 11. grupa. Skābekli saturošās fototrofās baktērijas. 12. grupa. Aerobās hemolitotrofās baktērijas un radniecīgi organismi. 13. grupa. Baktērijas, kas veido pumpurus un (vai) izaugumus. 14. grupa. Baktērijas ar apvalkiem. 15. grupa. Nefotosintētiskas slīdošās baktērijas, kas neveido augļķermeņus. 16. grupa. Slīdošās baktērijas, kas veido augļķermeņus. Nodaļā Firmicutes ietilpst: 17. grupa. Grampozitīvie koki. 18. grupa. Grampozitīvi nūjiņas un koki, kas veido endosporas. 19. grupa. Pareizas formas grampozitīvi nūjiņas, kas neveido sporas. 20. grupa. Neregulāras formas grampozitīvi nūjiņas, kas neveido sporas. Grupa 21. Mikobaktērijas. Grupas 22.–29. Aktinomicīti. Nodaļai Tenericutes pieder: 30. grupa. Mikoplazmas. Mendosicutes nodaļā ietilpst: 31. grupa. Metanogēni. 32. grupa. Sulfātus reducējošās arhebaktērijas. 33. grupa. Īpaši halofīlas arhebaktērijas (halobaktērijas). 34. grupa. Arhebaktērijas, kurām trūkst šūnu sienas. 35. grupa Īpaši termofīlas un hipertermofīlas sēru metabolizējošas arhebaktērijas. Noslēgumā jāuzsver, ka lielākā daļa dabiskajās kopienās esošo mikroorganismu joprojām ir jāizolē tīrkultūrās. Tiek uzskatīts, ka šobrīd ir iespējams kultivēt tikai 0,1% no kopējās mikrobu daudzveidības, un nav iespējams izaudzēt un identificēt atlikušos baktēriju pārstāvjus, lai gan jau ir izolēti un aprakstīti aptuveni 5 tūkstoši prokariotu sugu tīrkultūrā.