DNA-nukleotidin koostumus. Nukleotidit. Yhdiste. Rakenne. Nukleiinihappojen koostumus
Deoksiribonukleiinihapot (DNA) on makromolekyyli (yksi kolmesta päämolekyylistä, kaksi muuta ovat RNA:ta ja proteiineja), joka tarjoaa varastoinnin, siirtymisen sukupolvelta toiselle ja geneettisen ohjelman toteuttamisen elävien organismien kehitykselle ja toiminnalle. DNA sisältää tietoa erityyppisten RNA:iden ja proteiinien rakenteesta.
Eukaryoottisoluissa (eläimet, kasvit ja sienet) DNA:ta löytyy solun ytimestä osana kromosomeja sekä joissakin soluorganelleissa (mitokondrioissa ja plastideissa). Prokaryoottisten organismien (bakteerit ja arkit) soluissa pyöreä tai lineaarinen DNA-molekyyli, ns. nukleoidi, on kiinnittynyt sisältä solukalvoon. Heillä ja alemmilla eukaryooteilla (esimerkiksi hiivalla) on myös pieniä autonomisia, enimmäkseen pyöreitä DNA-molekyylejä, joita kutsutaan plasmideiksi. Lisäksi yksi- tai kaksijuosteiset DNA-molekyylit voivat muodostaa DNA:ta sisältävien virusten genomin.
Kemiallisesti katsoen DNA on pitkä polymeerinen molekyyli, joka koostuu toistuvista lohkoista - nukleotideista. Jokainen nukleotidi koostuu typpipitoisesta emäksestä, sokerista (deoksiriboosista) ja fosfaattiryhmästä. Ketjun nukleotidien väliset sidokset muodostuvat deoksiriboosista ja fosfaattiryhmästä (fosfodiesterisidokset). Suurimmassa osassa tapauksia (lukuun ottamatta joitakin yksijuosteista DNA:ta sisältäviä viruksia) DNA-makromolekyyli koostuu kahdesta ketjusta, jotka on suunnattu typpipitoisten emästen avulla toisiinsa. Tämä kaksijuosteinen molekyyli on kierteinen. Yleisesti ottaen DNA-molekyylin rakennetta kutsutaan "kaksoisheliksiksi".
DNA:n rakenteen purkaminen (1953) oli yksi biologian historian käännekohdista. Francis Crick, James Watson ja Maurice Wilkins saivat fysiologian tai lääketieteen Nobel-palkinnon vuonna 1962 erinomaisesta panoksestaan tähän löytöyn. Rosalind Franklin, joka sai röntgenkuvat, joita ilman Watson ja Crick eivät olisi voineet tehdä johtopäätöksiä DNA:n rakenne, kuoli vuonna 1958 syöpään, eikä Nobel-palkintoa jaeta kuoleman jälkeen.
Ribonukleiinihapot (RNA) on yksi kolmesta päämakromolekyylistä (kaksi muuta ovat DNA:ta ja proteiineja), joita löytyy kaikkien elävien organismien soluista.
Aivan kuten DNA (deoksiribonukleiinihappo), RNA koostuu pitkästä ketjusta, jossa jokaista linkkiä kutsutaan nukleotidiksi. Jokainen nukleotidi koostuu typpipitoisesta emäksestä, riboosisokerista ja fosfaattiryhmästä. Nukleotidisekvenssi sallii RNA:n koodata geneettistä tietoa. Kaikki solueliöt käyttävät RNA:ta (mRNA) ohjelmoimaan proteiinisynteesiä.
Solujen RNA muodostuu prosessissa, jota kutsutaan transkriptioksi, eli RNA:n synteesiin DNA-templaatilla, jonka suorittavat erityiset entsyymit - RNA-polymeraasit. Viesti-RNA:t (mRNA:t) osallistuvat sitten prosessiin, jota kutsutaan translaatioksi. Translaatio on proteiinin synteesi mRNA-templaatissa ribosomien osallistuessa. Muut RNA:t käyvät läpi kemiallisia modifikaatioita transkription jälkeen, ja sekundaaristen ja tertiääristen rakenteiden muodostumisen jälkeen ne suorittavat toimintoja, jotka riippuvat RNA:n tyypistä.
Yksijuosteisille RNA:ille on tunnusomaista useat avaruudelliset rakenteet, joissa jotkin saman ketjun nukleotideista ovat pariutuneet keskenään. Jotkut erittäin rakenteelliset RNA:t osallistuvat solun proteiinisynteesiin, esimerkiksi siirto-RNA:t tunnistavat kodoneja ja toimittavat vastaavat aminohapot proteiinisynteesikohtaan, kun taas ribosomaaliset RNA:t toimivat ribosomien rakenteellisena ja katalyyttisenä perustana.
RNA:n toiminnot nykyaikaisissa soluissa eivät kuitenkaan rajoitu niiden rooliin translaatiossa. Siten pienet tuman RNA:t osallistuvat eukaryoottisten lähetti-RNA:iden silmukointiin ja muihin prosesseihin.
Sen lisäksi, että RNA-molekyylit ovat osa joitakin entsyymejä (esimerkiksi telomeraasi), joillakin RNA:illa on oma entsymaattinen aktiivisuutensa: kyky tehdä katkoksia muihin RNA-molekyyleihin tai päinvastoin "liimata" kaksi RNA-fragmenttia. Tällaisia RNA:ita kutsutaan ribotsyymeiksi.
Useiden virusten genomit koostuvat RNA:sta, eli niissä sillä on rooli, jota DNA:lla on korkeammissa organismeissa. Perustuen RNA:n toimintojen monimuotoisuuteen solussa esitettiin hypoteesi, jonka mukaan RNA on ensimmäinen molekyyli, joka kykeni itse lisääntymään esibiologisissa järjestelmissä.
DNA:n ja RNA:n välillä on kolme pääeroa:
- 1. DNA sisältää sokerideoksiriboosin, RNA sisältää riboosia, jossa on ylimääräinen hydroksyyliryhmä deoksiriboosiin verrattuna. Tämä ryhmä lisää molekyylin hydrolyysin todennäköisyyttä, eli se vähentää RNA-molekyylin stabiilisuutta.
- 2. Adeniinille komplementaarinen nukleotidi RNA:ssa ei ole tymiini, kuten DNA:ssa, vaan urasiili on tymiinin metyloitumaton muoto.
- 3. DNA esiintyy kaksoiskierteen muodossa, joka koostuu kahdesta erillisestä molekyylistä. RNA-molekyylit ovat keskimäärin paljon lyhyempiä ja pääosin yksijuosteisia.
Biologisesti aktiivisten RNA-molekyylien, mukaan lukien tRNA, rRNA, snRNA ja muut molekyylit, jotka eivät koodaa proteiineja, rakenneanalyysi osoitti, että ne eivät koostu yhdestä pitkästä kierteestä, vaan useista lyhyistä helikseistä, jotka sijaitsevat lähellä toisiaan ja muodostavat jotain samanlaista kuin proteiinin tertiäärinen rakenne. Tämän seurauksena RNA voi katalysoida kemiallisia reaktioita, esimerkiksi ribosomin peptidyylitransferaasikeskus, joka osallistuu proteiinien peptidisidoksen muodostumiseen, koostuu kokonaan RNA:sta.
Vuoteen 1944 mennessä O. Avery ja hänen kollegansa K. McLeod ja M. McCarthy löysivät DNA:n muuntavan aktiivisuuden pneumokokeissa. Nämä kirjoittajat jatkoivat Griffithin työtä, joka kuvaili transformaatioilmiötä (perinnöllisten ominaisuuksien siirtymistä) bakteereissa. O. Avery, K. McLeod, M. McCarthy osoittivat, että kun proteiineja, polysakkarideja ja RNA:ta poistetaan, bakteerien transformaatio ei häiriinny, ja kun indusoiva aine altistuu deoksiribonukleaasientsyymille, transformoiva aktiivisuus katoaa.
Näissä kokeissa DNA-molekyylin geneettinen rooli osoitettiin ensimmäistä kertaa. Vuonna 1952 A. Hershey ja M. Chase vahvistivat DNA-molekyylin geneettisen roolin T2-bakteriofagilla tehdyissä kokeissa. Merkittämällä sen proteiinin radioaktiivisella rikillä ja sen DNA:n radioaktiivisella fosforilla, he infektoivat E. colin tällä bakteeriviruksella. Faagin jälkeläisistä löydettiin suuri määrä radioaktiivista fosforia ja vain pieniä määriä S:tä. Tästä seurasi, että DNA, ei faagin proteiini, tunkeutui bakteeriin ja siirtyi sitten replikaation jälkeen faagin jälkeläisiin. .
DNA-nukleotidin rakenne. Nukleotidien tyypit.
Nukleotidi DNA koostuu
Typpipitoinen emäs (4 tyyppiä DNA:ssa: adeniini, tymiini, sytosiini, guaniini)
Monosokerin deoksiriboosi
Fosforihappo
nukleotidimolekyyli koostuu kolmesta osasta - viiden hiilen sokerista, typpipitoisesta emäksestä ja fosforihaposta.
Sisältää sokeria nukleotidikoostumus, sisältää viisi hiiliatomia, eli se on pentoosi. Nukleotidissa olevan pentoosin tyypistä riippuen on olemassa kahdenlaisia nukleiinihappoja - ribonukleiinihappoja (RNA), jotka sisältävät riboosia, ja deoksiribonukleiinihapot (DNA), jotka sisältävät deoksiriboosia. Deoksiriboosissa OH-ryhmä toisessa hiiliatomissa korvataan H-atomilla, eli siinä on yksi happiatomi vähemmän kuin riboosissa.
Molemmissa nukleiinihappotyyppejä sisältää neljää erityyppistä emästä: niistä kaksi kuuluu puriinien luokkaan ja kaksi pyrimidiinien luokkaan. Renkaan sisältyvä typpi antaa näille yhdisteille pääluonteen. Puriineihin kuuluvat adeniini (A) ja guaniini (G) ja pyrimidiineihin kuuluvat sytosiini (C) ja tymiini (T) tai urasiili (U) (vastaavasti DNA:ssa tai RNA:ssa). Tymiini on kemiallisesti hyvin lähellä urasiilia (se on 5-metyyliurasiili, eli urasiili, jossa metyyliryhmä on viidennessä hiiliatomissa). Puriinimolekyylissä on kaksi rengasta, kun taas pyrimidiinimolekyylissä on yksi.
Nukleotidit on liitetty toisiinsa vahvalla kovalenttisella sidoksella yhden nukleotidin sokerin ja toisen fosforihapon kautta. Se käy ilmi polynukleotidiketju. Toisessa päässä on vapaa fosforihappo (5'-pää), toisessa on vapaa sokeri (3'-pää). (DNA-polymeraasi voi lisätä uusia nukleotideja vain 3'-päähän.)
Kaksi polynukleotidiketjua on liitetty toisiinsa heikoilla vetysidoksilla typpipitoisten emästen välillä. On 2 sääntöä:
komplementaarisuuden periaate: tymiini on aina adeniinin vastakohta, guaniini aina sytosiinin vastakohta (ne vastaavat toisiaan vetysidosten muodossa ja lukumäärässä - A:n ja G:n välillä on kaksi sidosta ja C:n ja G:n välillä 3).
antiparallelismin periaate: jossa yhdellä polynukleotidiketjulla on 5'-pää, toisella on 3'-pää ja päinvastoin.
Se käy ilmi kaksoisketju DNA.
Hän kiertyy kaksoiskierre, yksi kierteen kierros on pituus 3,4 nm, sisältää 10 paria nukleotideja. Typpipitoiset emäkset (geneettisen tiedon ylläpitäjät) ovat kierteen sisällä suojattuna.
Nukleiinihapot proteiinien tavoin ovat välttämättömiä elämälle. Ne edustavat kaikkien elävien organismien geneettistä materiaalia yksinkertaisimpiin viruksiin asti. Nimi "nukleiinihapot" kuvastaa sitä tosiasiaa, että ne sijaitsevat pääasiassa ytimessä (ydin - ydin). Nukleiinihappojen spesifisellä värjäyksellä ytimet näkyvät hyvin selvästi valomikroskoopissa.
DNA:n rakenteen selvittäminen(deoksiribonukleiinihappo) - yksi kahdesta olemassa olevasta nukleiinihapotyypistä - avasi uuden aikakauden biologiassa, koska se teki lopulta mahdolliseksi ymmärtää, kuinka elävät organismit säilyttävät elämänsä säätelemiseen tarvittavan tiedon ja kuinka ne välittävät tämän tiedon jälkeläisilleen. . Olemme jo edellä todenneet, että nukleiinihapot koostuvat monomeerisistä yksiköistä, joita kutsutaan nukleotideiksi. Äärimmäisen pitkät molekyylit - polynukleotidit - rakennetaan nukleotideista.
Polynukleotidien rakenteen ymmärtämiseksi on siksi ensin perehdyttävä miten rakennettuja nukleotideja.
Nukleotidit. Nukleotidien rakenne
nukleotidimolekyyli koostuu kolmesta osasta - viiden hiilen sokerista, typpipitoisesta emäksestä ja fosforipitoisesta emäksestä.
Sisältää sokeria nukleotidikoostumus, sisältää viisi hiiliatomia, eli se on pentoosi. Nukleotidissa olevan pentoosin tyypistä riippuen on olemassa kahdenlaisia nukleiinihappoja - ribonukleiinihappoja (RNA), jotka sisältävät riboosia, ja deoksiribonukleiinihapot (DNA), jotka sisältävät deoksiriboosia. Deoksiriboosissa OH-ryhmä toisessa hiiliatomissa korvataan H-atomilla, eli siinä on yksi happiatomi vähemmän kuin riboosissa.
Molemmissa nukleiinihappotyyppejä sisältää neljää erityyppistä emästä: niistä kaksi kuuluu puriinien luokkaan ja kaksi pyrimidiinien luokkaan. Renkaan sisältyvä typpi antaa näille yhdisteille pääluonteen. Puriineihin kuuluvat adeniini (A) ja guaniini (G) ja pyrimidiineihin kuuluvat sytosiini (C) ja tymiini (T) tai urasiili (U) (vastaavasti DNA:ssa tai RNA:ssa). Tymiini on kemiallisesti hyvin lähellä urasiilia (se on 5-metyyliurasiili, eli urasiili, jossa metyyliryhmä on viidennessä hiiliatomissa). Puriinimolekyylissä on kaksi rengasta, kun taas pyrimidiinimolekyylissä on yksi.
Säätiöt On tapana merkitä heidän nimensä ensimmäinen kirjain: A, G, T, U ja C.

Nukleiinihapot ovat happoja, koska niiden molekyyli sisältää fosforihappoa.
Kuvassa näkyy kuinka sokeri, emäs ja fosforihappo yhdistyvät muodostamaan nukleotidimolekyyli. Sokerin yhdistelmä emäksen kanssa tapahtuu vesimolekyylin vapautuessa, eli se on kondensaatioreaktio. Nukleotidin muodostamiseksi tarvitaan vielä yksi kondensaatioreaktio - sokerin ja fosforihapon välillä.
Sekalaiset nukleotidit poikkeavat toisistaan sokereiden ja niihin kuuluvien emästen luonteen suhteen.
Nukleotidien rooli kehossa ei rajoitu toimimaan nukleiinihappojen rakennuspalikoina; Jotkut tärkeät koentsyymit ovat myös nukleotideja. Näitä ovat esimerkiksi adenosiinitrifosfaatti (ATP), syklinen adenosiinimonofosfaatti (cAMP), koentsyymi A, ni(NAD), nikotiiniam(NADP) ja flaviiniadeniinidinukleotidi (FAD).
Nukleiinihapot ovat luonnollisia makromolekyyliyhdisteitä (polynukleotideja), joilla on valtava rooli perinnöllisen tiedon tallentamisessa ja välittämisessä elävissä organismeissa.
Nukleiinihappojen molekyylipaino voi vaihdella sadasta tuhansista kymmeniin miljardeihin. Ne löydettiin ja eristettiin soluytimistä jo 1800-luvulla, mutta niiden biologinen rooli selvitettiin vasta 1900-luvun jälkipuoliskolla.
Nukleotidin - nukleiinihappojen rakenneyksikön - koostumus sisältää kolme komponenttia:
1) typpipitoinen emäs - pyrimidiini tai puriini
Pyrimidiiniemäkset- pyrimidiinijohdannaiset, jotka ovat osa nukleiinihappoja:urasiili, tymiini, sytosiini.
–OH-ryhmän sisältäville emäksille on ominaista rakenteellisten isomeerien liikkuva tasapaino, joka johtuu protonin siirtymisestä hapesta typpeen ja päinvastoin:
Puriiniemäkset- puriinijohdannaiset, jotka ovat osa nukleiinihappoja: adeniini, guaniini.
Guaniini esiintyy kahtena rakenteellisena isomeerinä:
2) monosakkaridi
Riboosi ja 2-deoksiriboosi Termi "monosakkarideja" tarkoittaa viisi hiiliatomia sisältäviä monosakkarideja. Ne sisältyvät nukleiinihappojen koostumukseen syklisissä β-muodoissa:
3) fosforihappojäännös
DNA ja RNA
Riippuen siitä, minkä monosakkaridin polynukleotidin rakenneyksikkö sisältää - riboosi tai 2-deoksiriboosi, erottaa
· ribonukleiinihapot(RNA) ja
· deoksiribonukleiinihapot(DNA)
RNA:n pääjuoste (sokerifosfaatti) sisältää tähteitä riboosi ja DNA:ssa 2-deoksiriboosi.
DNA-makromolekyylien nukleotidiyksiköt voivat sisältää adeniini, guaniini, sytosiini ja tymiini. RNA:n koostumus eroaa sen sijaan tymiini esittää urasiili.
DNA:n molekyylipaino saavuttaa kymmeniä miljoonia amu. Nämä ovat pisimmät tunnetut makromolekyylit. RNA:n molekyylipaino on paljon pienempi (useista sadaista kymmeniin tuhansiin). DNA:ta löytyy pääasiassa solujen ytimistä, RNA:ta - solujen ribosomeista ja protoplasmasta.
Nukleiinihappojen rakennetta kuvattaessa otetaan huomioon makromolekyylien eri organisoitumistasot:ensisijainen ja toissijainen rakenne.
· Ensisijainen rakenne Nukleiinihapot on nukleotidikoostumus ja tietty sekvenssi nukleotidiyksiköitä polymeeriketjussa.
Esimerkiksi:
Lyhennetyssä yhden kirjaimen merkinnässä tämä rakenne on kirjoitettu muodossa
...- A - G - C -...
· Alla toissijainen rakenne nukleiinihapot ymmärtävät polynukleotidiketjujen spatiaalisesti järjestetyt muodot.
DNA:n toissijainen rakennekoostuu kahdesta rinnakkaisesta haarautumattomasta polynukleotidiketjusta, jotka on kierretty yhteisen akselin ympäri kaksoiskierteeksi.
Tällaista avaruudellista rakennetta ylläpitävät monet vetysidokset, jotka muodostuvat typpipitoisista emäksistä, jotka on suunnattu heliksin sisäänpäin.Vetysidoksia esiintyy yhden ketjun puriiniemäksen ja toisen ketjun pyrimidiiniemäksen välillä. Nämä emäkset muodostavat toisiaan täydentäviä pareja (lat. täydentävä- lisäys).
Vetysidosten muodostuminen komplementaaristen emäsparien välille johtuu niiden avaruudellisesta vastaavuudesta.
Pyrimidiiniemäs on komplementaarinen puriiniemäksen kanssa:
Vetysidokset muiden emäsparien välillä eivät salli niiden mahtua kaksoiskierrerakenteeseen. Täten,
TYMIINI (T) täydentää adeniinia (A),
SYTOSiini (C) on komplementaarinen GUANiinille (G).
Peruskomplementaarisuus määrääketjun täydentävyysDNA-molekyyleissä.
Polynukleotidiketjujen komplementaarisuus toimii kemiallisena perustana DNA:n päätehtävälle - perinnöllisten ominaisuuksien säilymiselle ja välittämiselle.
DNA:n kyky paitsi tallentaa, myös käyttää geneettistä tietoa määräytyy sen seuraavista ominaisuuksista:
DNA-molekyylit kykenevät replikoitumaan (kaksinkertaistumaan), ts. voi mahdollistaa muiden alkuperäisten kanssa identtisten DNA-molekyylien synteesin, koska emästen sekvenssi toisessa kaksoiskierteen ketjussa ohjaa niiden sijaintia toisessa ketjussa.
DNA-molekyylit voivat ohjata tietyn lajin organismeille spesifisten proteiinien synteesiä täysin tarkasti ja määrätietoisesti.
RNA:n toissijainen rakenne
Toisin kuin DNA, RNA-molekyylit koostuvat yhdestä polynukleotidiketjusta, eikä niillä ole tiukasti määriteltyä avaruudellista muotoa (RNA:n toissijainen rakenne riippuu niiden biologisista toiminnoista).
RNA:n päärooli on suora osallistuminen proteiinien biosynteesiin.
Tunnetaan kolmenlaisia solujen RNA:ta, jotka eroavat toisistaan sijainniltaan solussa, koostumukseltaan, koostaan ja ominaisuuksiltaan, jotka määräävät niiden erityisen roolin proteiinien makromolekyylien muodostumisessa:
informaatio (matriisi) RNA:t välittävät DNA:han koodattua tietoa proteiinin rakenteesta solun ytimestä ribosomeihin, joissa proteiinisynteesi suoritetaan;
kuljetus-RNA:t keräävät aminohappoja solun sytoplasmaan ja siirtävät ne ribosomiin; Tämän tyyppiset RNA-molekyylit "oppivat" lähetti-RNA-ketjun vastaavista osista, mitkä aminohapot tulisi osallistua proteiinisynteesiin;
Ribosomaaliset RNA:t tarjoavat tietyn rakenteen omaavaa proteiinisynteesiä lukemalla tietoa informaatio- (matriisi) RNA:sta.
ovat monimutkaisia monomeerejä, joista kootaan heteropolymeerimolekyylejä. DNA ja RNA. Vapaat nukleotidit osallistuvat elämän signaali- ja energiaprosesseihin. DNA-nukleotideillä ja RNA-nukleotideilla on yhteinen rakennesuunnitelma, mutta ne eroavat pentoosisokerin rakenteesta. DNA-nukleotidit käyttävät sokerideoksiriboosia, kun taas RNA-nukleotidit käyttävät riboosia.
Nukleotidin rakenne
Jokainen nukleotidi voidaan jakaa kolmeen osaan:
1. Hiilihydraatti on viisijäseninen pentoosisokeri (riboosi tai deoksiriboosi).
2. Fosforijäännös (fosfaatti) on fosforihapon jäännös.
3. Typpipitoinen emäs on yhdiste, jossa on monia typpiatomeja. Nukleiinihapoissa käytetään vain 5 tyyppiä typpipitoisia emäksiä: adeniini, tymiini, guaniini, sytosiini, urasiili. DNA:ssa on 4 tyyppiä: adeniini, tymiini, guaniini, sytosiini. RNA:ssa on myös 4 tyyppiä: Adeniini, Urasiili, Guaniini, Sytosiini.On helppo nähdä, että RNA:ssa tymiini on korvattu urasiililla DNA:han verrattuna.
Pentoosin (riboosin tai deoksiriboosin) yleinen rakennekaava, jonka molekyylit muodostavat nukleiinihappojen "rungon":
Jos X korvataan H:lla (X = H), saadaan deoksiribonukleosideja; jos X korvataan OH:lla (X = OH), saadaan ribonukleosideja. Jos korvaamme typpipitoisen emäksen (puriini tai pyrimidiini) R:n sijaan, saamme tietyn nukleotidin.
On tärkeää kiinnittää huomiota niihin hiiliatomien asemiin pentoosissa, jotka on merkitty 3" ja 5". Hiiliatomien numerointi alkaa yläosassa olevasta happiatomista ja kulkee myötäpäivään. Saadaan viimeinen hiiliatomi (5"), joka sijaitsee pentoosirenkaan ulkopuolella ja muodostaa, voisi sanoa, pentoosin "häntän", joten nukleotidiketjua rakentaessaan entsyymi pystyy kiinnittämään vain uuden nukleotidin hiileen 3" eikä mihinkään muuhun . Siksi nukleotidiketjun 5" päätä ei voida koskaan jatkaa; vain 3" päätä voidaan pidentää.



Vertaa RNA:n nukleotidia DNA:n nukleotidiin.
Yritä selvittää, mikä nukleotidi se on tässä esityksessä:

ATP - vapaa nukleotidi
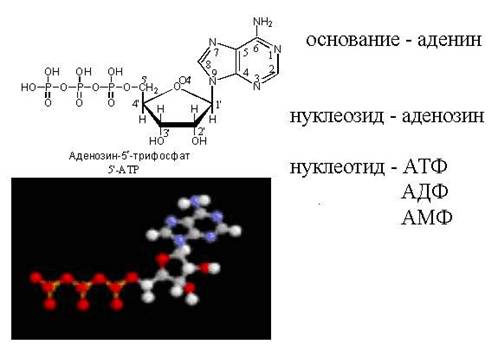
cAMP - "loopback" ATP-molekyyli

Nukleotidirakenteen kaavio

Huomaa, että aktivoidulla nukleotidilla, joka pystyy rakentamaan DNA- tai RNA-ketjun, on "trifosfaattipyrstö". Tämän "energialla kyllästetyn" hännän avulla se voi liittyä jo olemassa olevaan kasvavan nukleiinihapon ketjuun. Fosfaattipyrstö on hiilen 5 päällä, joten hiiliasema on jo fosfaattien varassa ja se on tarkoitettu kiinnitettäväksi. Mihin se kiinnittää? Vain hiileen kohdassa 3". Kiinnittyään tästä nukleotidista tulee itse kohde seuraavan nukleotidin kiinnittymiselle. "Vastaanottopuoli" tarjoaa hiilen asemassa 3", ja "saapuva puoli" tarttuu siihen fosfaattipyrstö, joka sijaitsee kohdassa 5". Yleensä ketju kasvaa 3":n puolelta.
DNA:n nukleotidiketjun pidentäminen
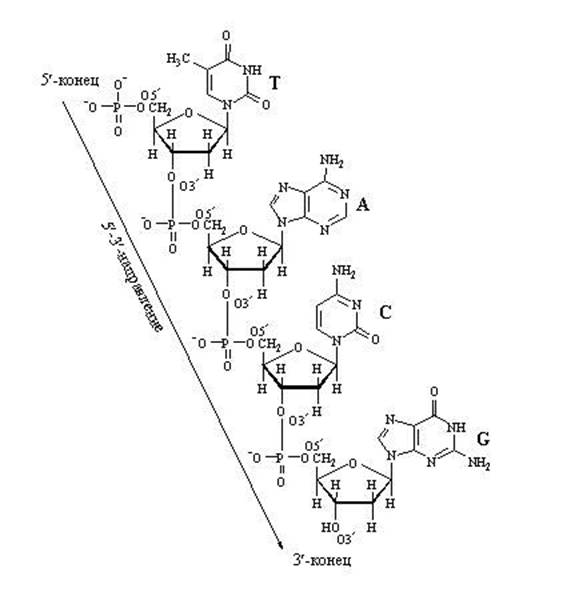
Nukleotidien välisten "pitkittäisten" sidosten aiheuttama ketjun kasvu voi mennä vain yhteen suuntaan: 5" ⇒ 3", koska Uusi nukleotidi voidaan lisätä vain ketjun 3'-päähän, ei 5'-päähän.
Nukleotidiparit, jotka on yhdistetty niiden typpipitoisten emästen "ristikkäisillä" komplementaarisilla sidoksilla
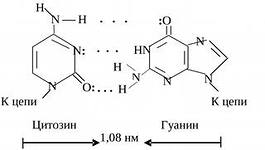
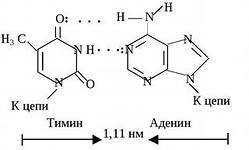
DNA:n kaksoiskierteen osa

Etsi merkkejä kahden DNA-juosteen antiparallelismista.
Etsi nukleotidiparit, joissa on kaksois- ja kolmoissidokset.