Mikro-organismien luokitus niiden päämuotojen mukaan. Bakteerit, niiden monimuotoisuus. Rakenne. Elinvoimaisuus. Perustermit: bakteerit
Bakteerien ravitsemus.
Ruokaa.
passiivinen diffuusio
Helpotettu diffuusio
aktiivinen kuljetus
Ensimmäisessä tapauksessa ravintoainemolekyyli muodostaa kompleksin periplasmisen tilaproteiinin kanssa, joka on vuorovaikutuksessa tietyn sytoplasmisen kalvopermeaasin kanssa. Energiasta riippuvan sytoplasmisen kalvon läpi tunkeutumisen jälkeen kompleksi "substraatti - periplasminen proteiini - permeaasi" dissosioituu substraattimolekyylin vapautuessa.
Aktiivisen kuljetuksen aikana kuljetettavan aineen kemiallisella modifikaatiolla tapahtumaketjuun kuuluu: (1) kalvoentsyymi-2:n fosforylaatio sytoplasmasta fosfoenolipyruvaatin vaikutuksesta; (2) fosforyloitujen entsyymi-2-substraattimolekyylien sitoutuminen sytoplasman kalvon pinnalle; (3) substraattimolekyylin energiariippuvainen kuljetus sytoplasmaan; (4) fosfaattiryhmän siirto substraattimolekyyliin; (5) "substraatti-entsyymi"-kompleksin dissosiaatio sytoplasmassa. Fosforylaatiosta johtuen substraattimolekyylit kerääntyvät solujen sytoplasmaan eivätkä pysty poistumaan niistä.
Bakteerien luokitus ruokatyypin mukaan.
Ravinteiden saannin kautta bakteerit luokitellaan holofyyttejä ja Holozoic. Holofyyttibakteerit (kreikasta. holos- täydellinen ja phyticos-kasveihin liittyvät) eivät pysty vapauttamaan ympäristöön substraatteja hajottavia entsyymejä, minkä seurauksena ne kuluttavat ravinteita yksinomaan liuenneessa, molekyylisessä muodossa. Holozoic bakteerit (kreikasta. holos- täydellinen ja zoikos- liittyvät eläimiin), päinvastoin, niillä on eksoentsyymien kompleksi, joka tarjoaa ulkoista ravintoa - substraattien hajoamista molekyyleiksi bakteerisolun ulkopuolella. Sen jälkeen ravintomolekyylit pääsevät holozoic-bakteerien sisälle.
Hiililähteen mukaan eristetty bakteereista autotrofit ja heterotrofit. Autotrofit (kreikasta. autot- itse, pokaali- ruoka) hiilidioksidia (CO 2) käytetään hiililähteenä, josta syntetisoidaan kaikki hiilipitoiset aineet. varten heterotrofit (kreikan kielestä geteros - toinen ja pokaali- ruoka) hiililähteet ovat erilaisia orgaanisia aineita molekyylimuodossa (hiilihydraatit, moniarvoiset alkoholit, aminohapot, rasvahapot). Korkein heterotrofian aste on luontainen prokaryooteille, jotka voivat elää vain muiden elävien solujen sisällä (esimerkiksi riketsia ja klamydia).
Typen lähteen mukaan prokaryootit jaetaan 3 ryhmään: 1) typpeä sitovat bakteerit (assimiloivat molekyylin typpeä ilmakehän ilmasta); 2) bakteerit, jotka kuluttavat epäorgaanista typpeä ammoniumsuoloista, nitriiteistä tai nitraateista; 3) bakteerit, jotka imevät orgaanisten yhdisteiden sisältämää typpeä (aminohapot, puriinit, pyrimidiinit jne.).
Energialähteen mukaan bakteerit jaetaan fototrofit ja kemotrofit. fototrofiset bakteerit , kuten kasvit, pystyvät käyttämään aurinkoenergiaa. Fototrofiset prokaryootit eivät aiheuta sairauksia ihmisille. Kemotrofiset bakteerit vastaanottaa energiaa redox-reaktioissa.
Elektronin luovuttajien luonteesta johtuen litotrofit(kreikasta. litos- kivi) ja organotrofit. klo litotrofit (kemolitotrofit ) epäorgaaniset aineet toimivat elektronien luovuttajina (H 2, H 2 S, NH 3, rikki, CO, Fe 2+ jne.). Elektronien luovuttajat organotrofit (kemoorganotrofit ) ovat orgaanisia yhdisteitä - hiilihydraatteja, aminohappoja jne.
Useimmilla ihmisille patogeenisilla bakteereilla on kemoorganotrofinen (kemoheterotrofinen) ravintotyyppi; kemolitotrofinen (kemoautotrofinen) tyyppi on vähemmän yleinen.
Kyky syntetisoida orgaanisia yhdisteitä kemotrofiset bakteerit luokitellaan prototrofit, auksotrofit ja hypotrofeja. prototrofiset bakteerit syntetisoi kaikki tarvittavat orgaaniset aineet glukoosista ja ammoniumsuoloista. Bakteereja kutsutaan auksotrofit jos ne eivät pysty syntetisoimaan mitään orgaanista ainetta mainituista yhdisteistä. Äärimmäistä metabolisen aktiivisuuden menetystä kutsutaan aliravitsemukseksi. hypotrofiset bakteerit tarjoavat elintärkeää toimintaansa järjestämällä uudelleen isännän rakenteita tai aineenvaihduntatuotteita.
Hiilen ja typen lisäksi rikki-, fosfori- ja metalli-ionit ovat välttämättömiä bakteerien täysimittaiselle elämälle. Rikin lähteitä ovat aminohapot (kysteiini, metioniini), vitamiinit, kofaktorit (biotiini, lipoiinihappo jne.), sulfaatit. Fosforin lähteitä ovat nukleiinihapot, fosfolipidit, fosfaatit. Riittävän korkeina pitoisuuksina bakteerit tarvitsevat magnesiumia, kaliumia, kalsiumia, rautaa; paljon pienempiä määriä - sinkki, mangaani, natrium, molybdeeni, kupari, nikkeli, koboltti.
kasvutekijöitä– Nämä ovat aineita, joita bakteerit eivät pysty itse syntetisoimaan, mutta ne tarvitsevat niitä kipeästi. Aminohapot, typpipitoiset emäkset, vitamiinit, rasvahapot, rautaporfyriinit ja muut yhdisteet voivat toimia kasvutekijöinä. Optimaalisten olosuhteiden luomiseksi bakteerien elintärkeälle aktiivisuudelle kasvutekijöitä on lisättävä ravintoalustaan.
Aineenvaihdunta, energian muuntaminen
A) Rakentava aineenvaihdunta.
Bakteeriravitsemuksen pakollinen vaihe on ravinteiden assimilaatio, eli niiden sisällyttäminen modifioidussa tai muunnetussa muodossa synteettisiin reaktioihin solukomponenttien ja -rakenteiden lisääntymiseksi.
Proteiinin aineenvaihdunta bakteereissa se voi edetä 3 vaiheessa: primaarinen proteiinien hajoaminen, sekundaarinen proteiinien hajoaminen ja proteiinisynteesi. Proteiinimolekyylien primaarisen hajoamisen peptoneiksi suorittaa eksoentsyymit - bakteerien ympäristöön vapauttamat eksoproteaasit. Toissijainen hajoaminen tapahtuu endoentsyymien (endoproteaasien) vaikutuksesta, joita kaikilla bakteereilla on. Tämä prosessi tapahtuu bakteerisolun sisällä ja koostuu peptidien hajoamisesta niiden muodostaviksi aminohapoiksi. Jälkimmäistä voidaan käyttää muuttumattomana tai altistaa kemiallisille muutoksille (deaminaatio, dekarboksylaatio jne.), jotka johtavat ammoniakin, indolin, rikkivedyn, ketohappojen, alkoholin, hiilidioksidin ym. ilmaantumiseen. Näiden yhdisteiden havaitseminen on diagnostinen merkitys bakteriologiassa.
Proteiinien pilkkoutumisreaktioiden ohella tapahtuu niiden synteesireaktioita. Jotkut bakteerit muodostavat proteiineja valmiista aminohapoista, jotka on saatu ulkoisen ravinnon tuloksena, kun taas toiset bakteerit syntetisoivat aminohappoja itsenäisesti yksinkertaisista typpeä ja hiiltä sisältävistä yhdisteistä. Aminohappojen synteesi voidaan suorittaa aminointi-, transaminaatio-, amidointi- ja karboksylaatioreaktioissa. Useimmat prokaryootit pystyvät syntetisoimaan kaikki solun proteiinit muodostavat aminohapot. Aminohappojen biosynteesin piirre on yleisten biosynteesireittien käyttö: trikarboksyylihapposykli, glykolyysi, oksidatiivinen pentoosi-fosfaattireitti jne. Aminohappojen synteesin pääasiallinen lähtöyhdiste on pyruvaatti ja fumaraatti.
hiilihydraattiaineenvaihduntaa se eroaa autotrofeista ja heterotrofeista (kaavio 1). Autotrofiset bakteerit syntetisoivat kaikki tarvittavat hiilihydraatit hiilidioksidista. Raaka-aineet hiilihydraattien muodostamiseksi heterotrofisissa bakteereissa voivat olla: (1) yhden, kahden ja kolmen hiilen yhdisteitä; ja (2) polysakkaridit (tärkkelys, glykogeeni, selluloosa). Jälkimmäisen pilkkomiseksi monilla heterotrofisilla bakteereilla on eksoentsyymejä (amylaasi, pektinaasi jne.), jotka hydrolysoivat polysakkarideja muodostaen glukoosia, maltoosia, fruktoosia jne.
Autotrofisissa bakteereissa Calvin-kierrossa hiilidioksidista muodostuu ribuloosifosfaatti-fosfori-glyseriinihappoa, joka sisältyy vastakkaiseen suuntaan tapahtuviin glykolyysireaktioihin. Käänteisen synteesin lopputuote on glukoosi.
Heterotrofiset bakteerit muodostavat glukoosia yhden, kahden ja kolmen hiilen yhdisteistä ja sisällyttävät ne myös käänteiseen glykolyysireaktioon. Koska jotkin glykolyysireaktiot ovat peruuttamattomia, heterotrofit ovat muodostaneet erityisiä entsymaattisia reaktioita, joiden avulla ne voivat ohittaa katabolisen reitin peruuttamattomat reaktiot.
Kun heterotrofiset bakteerit pilkkovat polysakkarideja, syntyneet disakkaridit pääsevät soluihin ja maltoosin, sakkaroosin ja laktoosin vaikutuksesta hydrolysoituvat ja hajoavat monosakkarideiksi, jotka sitten fermentoidaan tai osallistuvat sokerin interkonversion reaktioihin.
lipidien aineenvaihdunta. Sekä eksogeeniset lipidit että interstitiaalisen aineenvaihdunnan amfiboliitit voivat toimia lähtöaineina lipidien muodostukselle bakteereissa. Eksogeeniset lipidit altistuvat bakteerilipaaseille ja muille lipolyyttisille entsyymeille. Monet bakteerityypit imevät itseensä glyserolia, joka toimii muovimateriaalin ja energian lähteenä. Endogeeniset lipidisynteesin lähteet voivat olla asetyylikoentsyymi A, propionyyli-APB, malonyyli-APB (ACP - asetyylia kuljettava proteiini), fosfodioksiasetoni jne.
Parillisen hiiliatomimäärän rasvahappojen synteesin alkusubstraatti on asetyylikoentsyymi A, parittomien hiiliatomien rasvahapoille propionyyli-APB ja malonyyli-APB. Kaksoissidosten muodostuminen happomolekyylissä aerobisissa prokaryooteissa tapahtuu molekylaarisen hapen ja desaturaasin entsyymin osallistuessa. Anaerobisissa prokaryooteissa kaksoissidokset muodostuvat synteesin varhaisessa vaiheessa dehydraatioreaktion seurauksena. Fosfolipidien synteesin alkusubstraatti on fosfodioksiasetoni (glykolyyttisen reitin välituoteyhdiste), jonka pelkistyminen johtaa 3-fosforoglyserolin muodostumiseen. Sitten jälkimmäiseen lisätään 2 rasvahappojäännöstä kompleksin muodossa APB:n kanssa. Reaktiotuote on fosfatidihappo, jonka aktivoituminen CTP:llä ja sitä seuraava seriinin, inositolin, glyserolin tai muun yhdisteen fosfaattiryhmään liittyminen johtaa vastaavien fosfolipidien synteesiin.
Rasvahapoille auksotrofiset ja hypotrofiset mikro-organismit (esimerkiksi mykoplasmat) saavat ne valmiina isäntäsoluista tai ravintoalustasta.
Mononukleotidien vaihto. Puriini- ja pyrimidiinimononukleotidit ovat DNA:n ja RNA:n olennaisia komponentteja. Monet prokaryootit pystyvät sekä käyttämään ravintoalustaan sisältyviä valmiita puriini- ja pyrimidiiniemäksiä, niiden nukleosideja ja nukleotideja että syntetisoimaan niitä pienimolekyylisistä aineista. Bakteereilla on entsyymejä, jotka katalysoivat seuraavia eksogeenisten puriini- ja pyrimidiinijohdannaisten interkonversion vaiheita: typpiemäs - nukleosidi - nukleotidi (mono-di-trifosfaatti).
Puriini- ja pyrimidiinimononukleotidien synteesi de novo toteutetaan itsenäisillä tavoilla. Puriininukleotidien synteesin aikana peräkkäisten entsymaattisten reaktioiden seurauksena muodostuu inosiinihappoa, josta puriinirenkaan kemiallisilla modifikaatioilla syntetisoidaan adenyyli- (AMP) ja guanyylihappoja (GMP). Pyrimidiininukleotidien synteesi alkaa orotidyylihapon muodostumisella, jonka dekarboksylaatiosta saadaan uridyylihappoa (UMP). Jälkimmäisestä muodostuu UTP, jonka asylaatio johtaa CTP:n muodostumiseen.
Deoksiribonukleotidit muodostuvat vastaavien ribonukleotidien pelkistymisen seurauksena difosfaattien tai trifosfaattien tasolla. DNA-spesifisen nukleotidin, tymidyylihapon, synteesi tapahtuu deoksiuridyylihapon entsymaattisella metylaatiolla.
Ioninvaihto. Mineraaliyhdisteitä - ioneja, NH 3+, K+, Mg 2+, Fe 2+, SO 4 2-, PO 4 3- ja muita bakteereja saadaan ympäristöstä sekä vapaana että muuhun orgaaniseen liittyvässä tilassa. aineet. Kationit ja anionit kuljetetaan bakteerisoluun eri tavoin, jotka on kuvattu §:ssä 3. Ionien tunkeutumisnopeuteen bakteerisoluun vaikuttavat väliaineen pH ja itse mikro-organismien fysiologinen aktiivisuus.
B) Bakteerien hengitys (energian aineenvaihdunta).
Kaikki elämänprosessit ovat energiariippuvaisia, joten energian saanti on erittäin tärkeä osa prokaryoottien aineenvaihduntaa. He saavat energiaa anaerobisesta ja aerobisesta hengityksestä.
Hengitä, tai biologinen hapetus on katabolinen prosessi, jossa elektroneja siirtyy luovuttajaaineesta akseptoriaineeseen, johon liittyy energian kertymistä makroergisiin yhdisteisiin . Hengitys tapahtuu katabolisten reaktioiden prosessissa, jonka seurauksena monimutkaiset orgaaniset aineet halkeavat, luovuttavat energiaa ja muuttuvat yksinkertaisiksi yhdisteiksi. Makroergisiin aineisiin (ATP, GTP, UTP jne.) kertynyttä energiaa käytetään anabolisissa reaktioissa.
Hengitystavan mukaan mikro-organismit luokitellaan obligaatit (tiukat) aerobit, obligaatit anaerobit ja fakultatiiviset anaerobit.
pakolliset aerobit tarvitsevat vapaata happea. Orgaaniset yhdisteet (hiilihydraatit, rasvat, proteiinit) ovat elektronien luovuttajia ihmisille patogeenisissä kemoorganotrofisissa aerobeissa ja molekyylihappi on elektronien vastaanottaja. Energian varastointi ATP:n muodossa kemoorganotrofisissa aerobeissa tapahtuu elektronien luovuttajien oksidatiivisen fosforylaation aikana. Aerobeilla on sytokromeja (osallistuvat elektronien siirtoon) sekä entsyymejä (katalaasi, superoksididismutaasi, peroksidaasi), jotka inaktivoivat hengityksen aikana syntyviä myrkyllisiä happiradikaaleja. Superoksididismutaasi inaktivoi myrkyllisimmän metaboliitin, H 2 O 2:n superoksidiradikaalin. Entsyymi katalaasi muuttaa H 2 O 2:n H 2 O 2:ksi ja O 2:ksi.
Erityinen ryhmä aerobeja ovat mikroaerofiiliset bakteerit, jotka, vaikka ne tarvitsevat happea energiaksi, kasvavat paremmin lisääntyneen CO 2 -pitoisuuden myötä, esimerkiksi suvun bakteerit Kampylobakteeri ja Helicobacter.
pakolliset anaerobit eivät tarvitse vapaata happea, päinvastoin, jopa pieninä määrinä hapella on myrkyllinen vaikutus niihin. Elektronin luovuttajia ihmispatogeenisissa anaerobeissa-kemoorganotrofeissa ovat erilaisia orgaanisia yhdisteitä (pääasiassa hiilihydraatteja). Elektronien vastaanottaja kemoorganotrofisissa anaerobeissa on orgaanisia happea sisältäviä yhdisteitä - happoja tai ketoneja, eli elektronien vastaanottaja on orgaaniseen fragmenttiin liittyvä happi. Energian varastointi näissä prokaryooteissa tapahtuu substraattifosforylaation aikana. Pakollisilla anaerobeilla ei yleensä ole sytokromeja ja entsyymejä, jotka inaktivoivat happiradikaaleja (katalaasi- jaa).
Kemolitotrofisissa anaerobeissa, jotka eivät ole patogeenisiä ihmisille, elektronien vastaanottaja on epäorgaaniset happea sisältävät yhdisteet - nitraatit, sulfaatit, karbonaatit.
Erityinen ryhmä anaerobeja ovat aerotolerantit bakteerit, jotka pystyvät kasvamaan ilmakehän hapen läsnä ollessa, mutta eivät käytä sitä elektronien vastaanottajana (esimerkiksi maitohappobakteerit). Aerotolerantit katalaasi- ja sovat positiivisia.
Fakultatiiviset anaerobit pystyy esiintymään sekä happi- että hapettomassa ympäristössä. Niiden elektronin luovuttajat ovat orgaanisia aineita; elektronin vastaanottajat ovat ympäristöolosuhteista riippuen molekylaarisia tai happea sitoutuneita orgaanisiin ja epäorgaanisiin yhdisteisiin. Fakultatiiviset anaerobit voivat kerätä energiaa sekä oksidatiivisen että substraattifosforylaation aikana. Kuten aerobeissa, tässä bakteeriryhmässä on sytokromeja ja antioksidanttisia puolustusentsyymejä.
Pääsubstraatti energian saamiseksi ovat hiilihydraatit, jotka eri hengityksen kemoheterotrofisissa prokaryooteissa voivat kataboloitua asetyylikoentsyymi A:ksi ("aktivoitu etikkahappo"). Lipidit ja proteiinit voivat toimia energiasubstraatteina, koska asetyylikoentsyymi A on myös yksi niiden aineenvaihdunnan välituotteista (kaavio 2).
Hiilihydraattikatabolia kemoorganotrofisissa prokaryooteissa sisältää: (a) anaerobiset prosessit – glykolyysin, pentoosifosfaattireitin ja; (b) aerobinen prosessi – trikarboksyylihappokierto (Krebsin sykli). Anaerobisia prosesseja tapahtuu kaikissa prokaryooteissa, kun taas aerobinen prosessi on ominaista vain pakollisille aerobeille ja fakultatiivisille anaerobeille. Anaerobiset reitit perustuvat substraatin fosforylaatioon, kun taas aerobiset prosessit perustuvat oksidatiiviseen fosforylaatioon.
Käsitteiden määritelmä.
Sterilointi, desinfiointi ja antiseptiset aineet ovat olennaisia osa nykyaikaista lääketieteellistä ja erityisesti kirurgista toimintaa. Näiden menetelmien periaatteiden ja käytännön soveltamisen ymmärtäminen on välttämätöntä, koska monet mahdollisesti patogeeniset mikro-organismit voivat pysyä elävinä isännän ulkopuolella pitkiä aikoja, osoittavat suurta vastustuskykyä fysikaalisille ja kemiallisille desinfiointiaineille ja leviävät suhteellisen helposti ihmisestä toiseen.
Antiseptiset aineet- patogeenisten tai opportunististen mikro-organismien tuhoaminen tai kasvun estäminen kemiallisin menetelmin. Tätä termiä käytetään yleensä viittaamaan kemiallisen valmisteen ulkoiseen levittämiseen eläviin kudoksiin.
antiseptinen- aine, joka estää mikro-organismin kasvua tai tuhoaa sen (vaikuttamatta bakteeri-itiöihin). Termi koskee aineita, joita käytetään paikalliseen vaikutukseen eläviin kudoksiin.
Aseptinen tarkoittaa sepsiksen puuttumista, mutta yleisesti tätä termiä käytetään korostamaan elävien organismien puuttumista. Aseptiset menetelmät tarkoitetaan mitä tahansa menettelyä, joka on suunniteltu poistamaan eläviä organismeja ja estämään niiden aiheuttama uudelleensaastuminen. Nykyaikaiset kirurgiset ja mikrobiologiset tekniikat perustuvat aseptisiin toimenpiteisiin.
Biosidi- aine, joka tappaa kaikki elävät mikro-organismit, sekä patogeeniset että ei-patogeeniset, mukaan lukien itiöt.
Biostat- aine, joka estää mikro-organismien kasvun, mutta ei välttämättä tapa niitä.
Puhdistus- mikro-organismien poistaminen ilman kvantitatiivista määritystä. Tämä termi on suhteellinen; Lopullinen mikrobien poisto voidaan suorittaa steriloimalla tai desinfioimalla.
Desinfiointi- prosessi, joka vähentää tai eliminoi kaikki patogeenit itiöitä lukuun ottamatta.
Bakteerimyrkky- aine, joka tuhoaa mikro-organismeja, erityisesti taudinaiheuttajia. Bakteerimyrkky ei tuhoa itiöitä.
Sanaatio- menetelmä, jolla mikrobikontaminaatio vähennetään "turvalliselle" tasolle. Tätä menetelmää käytettiin aiemmin elottomien esineiden "puhdistamiseen".
Sterilointi- fyysisten tekijöiden ja (tai) kemikaalien käyttö kaikenlaisten mikrobielämän muotojen täydelliseen tuhoamiseen tai tuhoamiseen.
Sterilointi.
Sterilointi määritellään kaikkien mikro-organismien ja niiden itiöiden tuhoamiseksi tai poistamiseksi (suodatuksella). Sterilointi suoritetaan yleensä lämmöllä. Sterilointi, joka on yksi mikrobiologian laboratorion työskentelyn päivittäisistä rutiineista, on olennainen menetelmä varmistaa, että viljelmät, laitteet, välineet ja väliaineet tukevat vain välttämättömien mikro-organismien kasvua, kun taas muut mikrobit tuhoutuvat. On olemassa tällaisia sterilointityyppejä: kalsinointi polttimen liekissä, keittäminen, toiminta virtaavalla höyryllä, höyry paineella autoklaavissa, kuiva lämpö, pastörointi, tyndalointi, kemiallinen, kylmä (mekaaninen) sterilointi.
Sterilointimenetelmien valinta.
Sterilointimenetelmiä valittaessa on otettava huomioon seuraavat vaatimukset:
1. Aktiivisuus: bakteereja tappava, itiöitä tappava, tuberkuloosia tuhoava, fungisidinen ja virosidinen.
2. Toimenpiteen nopeus: sterilointi tulee suorittaa mahdollisimman nopeasti.
3. Läpäisevyys: Sterilointiaineiden on kyettävä tunkeutumaan pakkauksen läpi ja laitteen sisäpuolelle.
4. Yhteensopivuus: useita kertoja steriloitujen materiaalien rakenteessa tai toiminnassa ei saa tapahtua muutoksia.
5. Ei-myrkyllisyys: ihmisten terveydelle ja ympäristölle ei saa aiheutua uhkaa.
6. Orgaanisen materiaalin pysyvyys: steriloinnin tehokkuutta ei pitäisi heikentää orgaanisen materiaalin läsnä ollessa.
7. Muokattavuus: kyky käyttää suuria ja pieniä määriä steriloitua materiaalia.
8. Ajan hallinta: käsittelysykliä on ohjattava helposti ja tarkasti.
9. Hinta: kohtuulliset laitteet, asennus ja käyttö.
 Fyysiset sterilointilaitteet
Fyysiset sterilointilaitteet
märkä lämmin, höyryautoklavoinnin aikana muodostuva aine on tärkein kliinisissä mikrobiologian laboratorioissa käytetty sterilointiaine. Autoklaaveja käytetään viljelyalustojen, tulenkestävän materiaalin sterilointiin ja tartuntajätteen käsittelyyn. Höyrysterilointilaite tai autoklaavi on eristetty paineistettu kammio, joka käyttää kyllästettyä höyryä korkeiden lämpötilojen tuottamiseen (kuva 1). Ilma poistetaan kammiosta massasiirtymällä tai tyhjiöllä. Yleisimmin käytetyt autoklaavit painon mukaan vaihdetuilla. Kammioon johdetaan kevyempää höyryä raskaamman ilman syrjäyttämiseksi. Lyhytaikainen altistuminen paineistetulle höyrylle voi tuhota bakteeri-itiöitä. Viljelyalustojen ja muiden materiaalien rutiininomaisessa steriloinnissa altistusaika on 15 minuuttia 121 °C:ssa ja 1,5 kg:n paineessa 1 neliösenttimetriä kohti. Tartuntajätteen osalta altistusaika pidennetään 30-60 minuuttiin. Oikean ajan ja lämpötilan lisäksi suora kosketus höyryn kanssa on erittäin tärkeää steriloinnissa. Tarttuvaa materiaalia käsiteltäessä on varmistettava, että jätteeseen pääsee mahdollisimman paljon höyryä. Tällainen materiaali on käsiteltävä 132 ºC:n lämpötilassa. Antineoplasteja, myrkyllisiä kemikaaleja ja radioisotooppeja, jotka eivät välttämättä hajoa, eikä epävakaita kemikaaleja ei tule autoklavoida, koska ne voivat haihtua ja levitä kammion läpi altistuessaan lämmölle.
 Kuivakuumsterilointi käytetään materiaaleihin, joita ei voida höyrysteriloida vaurioitumisen mahdollisuuden tai materiaalin höyrynpitämättömyyden vuoksi. Kuiva lämpö on vähemmän tehokasta kuin kostea lämpö ja vaatii pidemmät altistusajat ja korkeammat lämpötilat. Kuivalämmöllä sterilointi suoritetaan yleensä kuivalämpökaapissa (kuva 2). Kuivalämpösteriloinnin mekanismi on oksidatiivinen prosessi. Esimerkkejä materiaaleista, joissa käytetään kuivalämpösterilointia, ovat öljyt, jauheet, terävät instrumentit ja lasitavarat. Vaihtoehtoisina menetelminä tartuntajätteen käsittelyssä käytetään kuivaa lämpöä tai lämpöinaktivointi-sterilointia.
Kuivakuumsterilointi käytetään materiaaleihin, joita ei voida höyrysteriloida vaurioitumisen mahdollisuuden tai materiaalin höyrynpitämättömyyden vuoksi. Kuiva lämpö on vähemmän tehokasta kuin kostea lämpö ja vaatii pidemmät altistusajat ja korkeammat lämpötilat. Kuivalämmöllä sterilointi suoritetaan yleensä kuivalämpökaapissa (kuva 2). Kuivalämpösteriloinnin mekanismi on oksidatiivinen prosessi. Esimerkkejä materiaaleista, joissa käytetään kuivalämpösterilointia, ovat öljyt, jauheet, terävät instrumentit ja lasitavarat. Vaihtoehtoisina menetelminä tartuntajätteen käsittelyssä käytetään kuivaa lämpöä tai lämpöinaktivointi-sterilointia.
Pastörointi tuhoaa taudinaiheuttajat kuumentamalla ainetta nopeasti 71,1 °C:seen 15 sekunniksi, minkä jälkeen jäähdytetään nopeasti. Pastörointi ei ole sterilointia, koska kaikki mikro-organismit eivät ole sille herkkiä. Tämä menetelmä eliminoi sairauksien, kuten ruoansulatuskanavan tuberkuloosin ja Q-kuumeen, leviämisen elintarviketeitse.
Tyndalisaatio on ajoittainen lämpösterilointimenetelmä, jolla voidaan tappaa kaikki bakteerit liuoksissa. Koska kasvavat bakteerit tappavat helposti lyhyillä keitoilla (5 kertaa 1 tunnin sisällä 5 minuutin ajan), liuoksen on annettava seistä tietty aika, ennen kuin lämpö häiritsee itiöiden kypsymistä merkittävästi. niiden lämmönkestävyydestä.
Suodatus on prosessi, jolla poistetaan mikrobit ja mikroskooppiset hiukkaset liuoksista, ilmasta ja muista kaasuista. Yleisin suodattamalla sterilisoinnin käyttö laboratoriossa on diagnostiikan, viljelyalustojen, kudosviljelyalustojen, seerumien, seerumikomponentteja sisältävien liuosten hoitoon. Toinen yleinen suodatuksen sovellus on ilman ja kaasujen sterilointi. Muoviset tai paperikalvosuodattimet, jotka erottuvat huokoshalkaisijaltaan (noin 12 - 0,22 µm) ja joita käytetään mekaaniseen erotukseen, toimivat myös mikrobien keräämiseen nesteistä mikroskooppista tutkimusta tai viljelyä varten suoraan suodattimella, kun se asetetaan pinnalle. liotettu ravintoalusta.
ultraviolettisäteilyä on eräänlainen sähkömagneettinen aaltosäteily, joka vaikuttaa solujen nukleiinihappoihin. Mikro-organismit ovat erittäin herkkiä ultraviolettisäteiden vaikutukselle, jonka aallonpituus on 254 nm. Ultraviolettivaloa käytetään yleisimmin ilmassa tai pinnoilla olevien mikro-organismien tappamiseen. Muita käyttökohteita ovat tiettyjen kemikaalien ja muovien kylmästerilointi lääkesovelluksissa, seerumisterilointi soluviljelmille ja veden desinfiointi. Ultraviolettisäteilyn merkittävä haitta sterilointilaitteena on sen kyvyttömyys tunkeutua materiaaleihin.
ionisoiva säteily sähkömagneettisessa spektrissä sillä on tappava vaikutus mikro-organismeihin. Tämä spektri sisältää mikroaallot, y-säteet, röntgensäteet ja elektronivirran. Ionisoivan säteilyn tappava vaikutus johtuu suorasta vaikutuksesta kohdemolekyyliin, minkä seurauksena energia siirtyy molekyyliin; ja epäsuoran toiminnan vuoksi - radikaalien diffuusio.
ultraäänienergiaa alhaisella taajuudella inaktivoi mikro-organismeja vesiliuoksissa. Sonikoinnin fyysinen vaikutus johtuu kavitaatiosta. Ultraäänipuhdistimia ja muita laitteita käytetään usein instrumenttien puhdistamiseen, mutta niitä ei pidetä steriloijina. Ultraäänen ja kemiallisen käsittelyn yhdistelmä tappaa kuitenkin mikro-organismeja.
Kemialliset sterilointilaitteet
2 % glutaraldehydi Nestemäisenä kemiallisena sterilointiaineena sitä käytettiin aiemmin laajalti sellaisten lääketieteellisten ja kirurgisten materiaalien käsittelyyn, joita ei voida steriloida kuumentamalla tai säteilyttämällä. Glutaraldehydiä käytetään myös rokotteiden valmistuksessa.
Desinfiointi.
Desinfiointi voidaan suorittaa kemiallisilla menetelmillä tai keittämällä. Keittäminen on tehokas menetelmä instrumenttien, kuten neulojen ja ruiskujen, desinfiointiin, jos autoklaavia ei ole saatavilla. Esipuhdistettuja lääketieteellisiä instrumentteja tulee keittää 20 minuuttia. Kemiallista desinfiointia käytetään lämpöherkille laitteille, jotka voivat vaurioitua korkeissa lämpötiloissa. Kemiallisia desinfiointiaineita, kuten kloorikomponentteja, etyyli- ja isopropyylialkoholia, kvaternäärisiä ammoniumkomponentteja ja glutaraldehydiä, käytetään laajalti.
Kemialliset desinfiointiaineet.
Alkoholi (etyyli ja isopropyyli), liuotettuna veteen 60-85 %:n pitoisuuteen, on erittäin tehokas desinfioinnissa. Alkoholit ovat bakteereja, sieniä ja tuberkuloosia tappavia, mutta eivät vaikuta itiöihin. Etyylialkoholilla on laajempi virosidinen vaikutus kirjo kuin isopropyylialkoholilla, joten se on tehokkaampi lipofiilisiä ja hydrofiilisiä viruksia vastaan.
Ratkaisu 37% formaldehydi, jota kutsutaan formaliiniksi, voidaan käyttää sterilointiaineena, kun taas sen pitoisuudet 3-8 % voidaan käyttää desinfiointiaineina.
Fenoli puhtaassa muodossaan ei käytetä desinfiointiaineena johtuen sen myrkyllisyydestä, kyvystä aiheuttaa kasvainten kehittymistä ja korroosiota. Fenolijohdannaisia, joissa funktionaalinen ryhmä (kloori, bromi, alkyyli, bentsyyli, fenyyli, amyyli) korvaa yhden aromaattisen renkaan vetyatomin, käytetään laajalti desinfiointiaineina. Tämä korvaaminen vähentää fenolin haittoja. Fenolin komponentit tappavat mikrobeja entsyymijärjestelmien inaktivoitumisen, proteiinien saostumisen ja soluseinän ja kalvon hajoamisen vuoksi. Yleensä käytetään 2-5 %:n pitoisuuksia, pienemmät pitoisuudet vaativat pidemmän altistuksen.
Halogeenit. Laboratoriokäytännössä desinfiointiin käytetään vain klooria ja jodia. Koska kloori on voimakas hapetin, sen uskotaan tappavan mikrobeja hapettumalla. Uskotaan, että jodi tappaa mikro-organismeja reagoimalla aminohappojen N-H- ja S-H-ryhmien sekä tyrosiini-aminohapon fenoliryhmän ja tyydyttymättömien rasvahappojen hiili-hiili-kaksoissidosten kanssa. Perinteisessä käsittelyssä ruiskutetaan 2-5 % formaldehydiliuosta höyryn läsnä ollessa 60-80 ºC lämpötilassa.
Antiseptiset aineet.
Antiseptisia aineita löytyy mikrobiologisista laboratorioista, pääasiassa käsienpesuun käytettävistä aineista. Tapauksissa, joissa lääkintähenkilöstö antaa ensiapua potilaille antibakteerisia aineita sisältävillä aineilla, tämä vähentää sairaalainfektioiden määrää. Yleisimmät käsienpesussa esiintyvät kemikaalit ovat alkoholit, klooriheksidiiniglukonaatti, jodoforit, klooriksylenoli ja triklosaani.
Perinteisiä jätteiden ja roskien käsittelymenetelmiä ovat poltto ja höyrysterilointi.
Palaa on paras tapa käsitellä jätteitä ja roskia. Tämä menetelmä tekee jätteestä tarttumattoman ja muuttaa myös sen muotoa ja kokoa. Sterilointi on tehokas jätteenkäsittelymenetelmä, mutta se ei muuta muotoaan. Höyrysterilointi autoklaavissa 121 °C:ssa vähintään 15 minuutin ajan tappaa kaikki mikrobielämän muodot, mukaan lukien suuret bakteeri-itiöt. Tämän tyyppinen täydellinen sterilointi voidaan tehdä myös kuivalla lämmöllä 160-170 ºC:ssa 2-4 tunnin ajan. On kuitenkin varmistettava, että kuiva lämpö on kosketuksissa steriloitavaan materiaaliin. Siksi nestettä sisältävät pullot on suljettava löyhästi korkilla tai vanupuikolla, jotta höyry ja lämpö voivat vaihtua pulloissa olevan ilman kanssa. Biovaaralliset jätteet sisältävät säiliöt tulee sitoa tiukasti. Steriloitu biologisesti vaarallinen materiaali on suljettava asianmukaisesti merkittyihin astioihin.
Höyrysterilointi (autoklaavi). Tartuntajätteet katsotaan dekontaminoituneiksi, kun vegetatiivisten bakteerien, sienten, mykobakteerien ja lipidejä sisältävien virusten määrä vähenee 6 lg-kertaisesti ja bakteerien endosporien määrä 4 lg-kertaisesti.
Bakteerien ravitsemus.
Ruokaa. Bakteerisolun ravinnon alla tulee ymmärtää muovimateriaalin ja energian absorptio- ja assimilaatioprosessi transformatiivisten reaktioiden seurauksena . Prokaryoottien ravitsemustyypit ovat monimutkaisia ja monipuolisia. Ne vaihtelevat riippuen tavasta, jolla ravinteet pääsevät bakteerisoluun, hiilen ja typen lähteistä, energian saantitavasta ja elektronien luovuttajien luonteesta.
ravinteiden kuljetus soluun voidaan suorittaa kolmella mekanismilla: passiivinen diffuusio, helpotettu diffuusio ja aktiivinen kuljetus.
passiivinen diffuusio on epäspesifinen energiariippuvainen prosessi, joka suoritetaan aineiden pitoisuusgradienttia pitkin (aine korkeamman pitoisuuden omaavasta ympäristöstä passiivisesti, osmoosin lakien mukaan, tulee ympäristöön, jonka pitoisuus on pienempi). Passiivisen diffuusion avulla bakteerisoluun pääsee rajallinen määrä aineita, joitain ioneja, monosakkarideja. Aineiden siirtonopeus passiivisen diffuusion aikana on merkityksetön ja riippuu kuljetettavien molekyylien lipofiilisyydestä ja koosta.
Helpotettu diffuusio on haihtumaton aineiden kuljetus pitoisuusgradienttia pitkin permeaasientsyymien avulla. Permeaasit ovat spesifisiä kalvoproteiineja, jotka helpottavat aineiden kulkeutumista sytoplasman kalvon läpi. Permeaasi kiinnittää itseensä siirretyn aineen molekyylin, jonka kanssa se voittaa sytoplasmisen kalvon, minkä jälkeen "aine-permeaasi" -kompleksi dissosioituu. Vapautunutta permeaasia käytetään muiden molekyylien johtamiseen. Prokaryooteissa vain glyseroli pääsee soluun helpotetun diffuusion kautta. Tässä tapauksessa glyserolin solunsisäinen pitoisuus vastaa solun ulkopuolista pitoisuutta. Helpotettu diffuusio on tyypillisin eukaryoottisille mikro-organismeille.
aktiivinen kuljetus- tämä on energiariippuvaista aineiden siirtymistä soluun pitoisuusgradienttia vastaan tiettyjen entsyymien avulla. Suurin osa aineista (ionit, hiilihydraatit, aminohapot, lipidit jne.) pääsee bakteerisoluun aktiivisen kuljetuksen kautta. Aktiivinen kuljetus voi tapahtua: (1) ilman kuljetettavan aineen kemiallista modifiointia; (2) kemiallisesti modifioituna.
Bakteerit ovat solurakenteen prokaryoottisia mikro-organismeja. Niiden koot ovat 0,1 - 30 mikronia. Mikrobit ovat erittäin yleisiä. Ne elävät maaperässä, ilmassa, vedessä, lumessa ja jopa kuumissa lähteissä, eläinten kehossa sekä elävien organismien sisällä, mukaan lukien ihmiskeho.
Bakteerien jakautuminen lajeihin perustuu useisiin kriteereihin, joista useimmiten huomioidaan mikro-organismien muoto ja niiden tilajakauma. Joten solujen muodon mukaan bakteerit jaetaan:
Coci - mikro-, diplo-, strepto-, stafylokokit sekä sarkiinit;
sauvan muotoinen - monobakteerit, diplobakteerit ja streptobakteerit;
Kierteiset lajit - vibriot ja spirokeetat.
Burgey's-determinantti systematisoi kaikki tunnetut bakteerit käytännön bakteriologiassa laajimman levinneisyyden löytäneiden bakteerien tunnistamisen periaatteiden mukaisesti perustuen soluseinän rakenteen eroihin ja suhteessa Gram-värjäytymiseen. Bakteerien kuvaus on annettu ryhmittäin (osioittain), joihin kuuluvat perheet, suvut ja lajit; joissakin tapauksissa ryhmät sisältävät luokkia ja tilauksia. Ihmisille patogeeniset bakteerit sisältyvät pieneen määrään ryhmiä.
Avain erottaa neljä bakteerien pääluokkaa -
Gracillicutes [sanasta lat. gracilis, siro, ohut, + cutis, skin] - lajit, joilla on ohut soluseinä, värjäytynyt gram negatiivinen;
firmicutes [lat. flrmus, vahva, + cutis, skin] - bakteerit, joilla on paksu soluseinä, värjäytyminen grampositiivinen;
Tenericutes [sanasta lat. kireä, hellävarainen, + cutis, iho] - bakteerit, joilta puuttuu soluseinä(mykoplasmat ja muut Mollicutes-luokan jäsenet)
Mendosicutes [lat. mendosus, epäsäännöllinen, + cutis, iho] - arkebakteerit (metaania ja sulfaattia pelkistävät, halofiiliset, termofiiliset ja arkebakteerit, joissa ei ole soluseinää).
Ryhmä 2 Burgeyn määräävä tekijä. Aerobiset ja mikroaerofiiliset liikkuvat kierteiset ja kaarevat Gram-negatiiviset bakteerit. Ihmisille patogeeniset lajit sisältyvät Campylobacter-, Helicobacters Spirillum -sukuihin.
Bergeyn determinantin ryhmä 3. Ei-liikkuvat (harvoin liikkuvat) Gram-negatiiviset bakteerit. Ei sisällä patogeenisiä lajeja.
Burgeyn determinantin ryhmä 4. Gram-negatiiviset aerobiset ja mikroaerofiiliset sauvat ja kokit. Ihmisille patogeeniset lajit kuuluvat Legionellaceae-, Neisseriaceae- ja Pseudomonada-ceae-heimoihin, ryhmään kuuluvat myös patogeeniset ja opportunistiset bakteerit suvuista Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Francisella, Kingella ja Moraxella.
Bergeyn determinantin ryhmä 5. Fakultatiiviset anaerobiset Gram-negatiiviset sauvat. Ryhmän muodostaa kolme perhettä - Enterobacteriaceae, Vibrionaceae ja Pasteurellaceae, joista jokainen sisältää patogeenisiä lajeja sekä patogeenisiä ja opportunistisia bakteereja suvuista Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella ja Streptobacillus.
Bergeyn determinantin ryhmä 6. Gram-negatiiviset anaerobiset suorat, kaarevat ja spiraaliset bakteerit. Patogeeniset ja opportunistiset lajit sisältyvät Bacteroides-, Fusobacterium-, Porphoromonas- ja Prevotelta-sukuihin.
Bergeyn determinantin ryhmä 7. Bakteerit, jotka suorittavat sulfaatin tai rikin dissimilaatiopelkistyksen Ei sisällä patogeenisiä lajeja.
Bergeyn determinantin ryhmä 8. Anaerobiset gramnegatiiviset kokit. Sisältää Veillonella-suvun opportunistisia bakteereja.
Bergeyn determinantin ryhmä 9. Rickettsia ja klamydia. Kolme perhettä - Rickettsiaceae, Bartonellaceae ja Chlamydiaceae, joista jokainen sisältää ihmisille patogeenisiä lajeja.
Burgeyn oppaan ryhmiin 10 ja 11 kuuluvat anoksi- ja happipitoiset fototrofiset bakteerit, jotka eivät ole patogeenisiä ihmisille.
Burgeyn determinantin ryhmä 12. Aerobiset kemolitotrofiset bakteerit ja niihin liittyvät organismit. Siinä yhdistyvät rikkiä, rautaa ja mangaania hapettavat ja nitrifioivat bakteerit, jotka eivät aiheuta vahinkoa ihmisille.
Burgeyn oppaan ryhmiin 13 ja 14 kuuluvat orastavat ja/tai uloskasvubakteerit ja tuppia muodostavat bakteerit. Edustaa vapaasti eläviä lajeja, jotka eivät ole patogeenisiä ihmisille;
Burgeyn oppaan ryhmät 15 ja 16 yhdistävät liukubakteerit, jotka eivät muodosta hedelmäkappaleita ja muodostavat niitä. Ryhmät eivät sisällä ihmisille patogeenisiä lajeja.
Burgeyn determinantin ryhmä 17. Gram-positiiviset kokit. Sisältää opportunistiset lajit suvuista Enterococcus Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus.
Burgeyn determinantin ryhmä 18. Itiöitä muodostavat grampositiiviset sauvat ja kokit. Sisältää patogeeniset, ehdollisesti patogeeniset sauvat suvuista Clostridium ja Bacillus.
Burgeyn determinantin ryhmä 19. Itiöitä muodostavat säännöllisen muotoiset grampositiiviset sauvat. Mukaan lukien opportunistiset lajit suvuista Erysipelothrix ja Listeria.
Burgeyn determinantin ryhmä 20. Epäsäännöllisen muotoisia itiöitä muodostavia Gram-positiivisia sauvoja. Ryhmään kuuluvat patogeeniset ja opportunistiset lajit suvuista Actinomyces, Corynebacterium Gardnerella, Mobiluncus jne.
Burgeyn determinantin ryhmä 21. Mykobakteerit. Sisältää ainoan Mycobacterium-suvun, joka yhdistää patogeeniset ja opportunistiset lajit.
Ryhmät 22-29. Actinomycetes. Lukuisista lajeista vain Gordona-, Nocardia-, Rhodococcus-, Tsukamurella-, Jonesia-, Oerskovi- ja Terrabacter-sukujen nokardiomuotoiset aktinomykeetit (ryhmä 22) kykenevät aiheuttamaan vaurioita ihmisissä.
Burgeyn determinantin ryhmä 30. Mykoplasmat. Acholeplasma-, Mycoplasma- ja Ureaplasma-sukuun kuuluvat lajit ovat patogeenisiä ihmisille.
Muut Bergeyn determinanttien ryhmät - metanogeeniset bakteerit (31), sulfaattia pelkistävät bakteerit (32 erittäin halofiilistä aerobista arkebakteeria (33), arkkibakteerit, joissa ei ole soluseinämää (34), äärimmäiset termofiilit ja hypertermofiilit, rikkiä metaboloivat bakteerit (35) - eivät sisältää ihmisille patogeenisiä lajeja.
Bakteerit ovat vanhin maapallolla tällä hetkellä olemassa oleva organismiryhmä. Ensimmäiset bakteerit ilmestyivät luultavasti yli 3,5 miljardia vuotta sitten ja olivat lähes miljardin vuoden ajan ainoat elävät olennot planeetallamme. Koska nämä olivat ensimmäisiä villieläinten edustajia, heidän ruumiillaan oli primitiivinen rakenne.
Ajan myötä niiden rakenne muuttui monimutkaisemmaksi, mutta nykyäänkin bakteereja pidetään primitiivisimpinä yksisoluisina organismeina. Mielenkiintoista on, että jotkut bakteerit säilyttävät edelleen muinaisten esi-isiensä primitiiviset piirteet. Tämä havaitaan bakteereissa, jotka elävät kuumissa rikkilähteissä ja hapettomissa lieteissä säiliöiden pohjalla.
Useimmat bakteerit ovat värittömiä. Vain harvat ovat väriltään violetteja tai vihreitä. Mutta monien bakteerien pesäkkeillä on kirkas väri, mikä johtuu värillisen aineen vapautumisesta ympäristöön tai solujen pigmentaatiosta.
Bakteerimaailman löytäjä oli Anthony Leeuwenhoek, 1600-luvun hollantilainen luonnontieteilijä, joka loi ensimmäisenä täydellisen suurennuslasimikroskoopin, joka suurentaa esineitä 160-270 kertaa.
Bakteerit luokitellaan prokaryootiksi ja erotetaan erilliseen valtakuntaan - Bakteerit.
kehonmuoto
Bakteerit ovat lukuisia ja erilaisia organismeja. Ne eroavat muodoltaan.
| bakteerin nimi | Bakteerin muoto | Bakteerikuva |
| cocci | pallomainen | |
| Basilli |  | sauvan muotoinen |
| Vibrio | kaareva pilkku | |
| Spirillum |  | Kierre |
| streptokokit |  | Cocci-ketju |
| Stafylokokit |  | Kokkirypäleitä |
| diplokokit | Kaksi pyöreää bakteeria yhden limaisen kapselin sisällä |
Kuljetustavat
Bakteerien joukossa on liikkuvia ja liikkumattomia muotoja. Liikkuvat liikkuvat aaltomaisten supistusten tai erityisestä flagelliiniproteiinista koostuvien flagellan (kierrettyjen kierteisten lankojen) avulla. Niissä voi olla yksi tai useampi siima. Ne sijaitsevat joissakin bakteereissa solun toisessa päässä, toisissa - kahdessa tai koko pinnalla.

Mutta liike on luontaista myös monille muille bakteereille, joilla ei ole siimat. Joten ulkopuolelta liman peittämät bakteerit pystyvät liukumaan.
Joidenkin vesi- ja maabakteerien sytoplasmassa on kaasuvakuoleja, joissa ei ole flagellaa. Solussa voi olla 40-60 vakuolia. Jokainen niistä on täytetty kaasulla (oletettavasti typellä). Säätelemällä tyhjiöissä olevan kaasun määrää vesibakteerit voivat vajota vesipatsaan tai nousta sen pinnalle, kun taas maaperän bakteerit voivat liikkua maaperän kapillaareissa.
Habitat
Organisaation yksinkertaisuuden ja vaatimattomuuden vuoksi bakteerit ovat laajalle levinneitä luonnossa. Bakteereja löytyy kaikkialta: pisarasta jopa puhtainta lähdevettä, maaperän jyväissä, ilmassa, kivillä, napalumissa, aavikon hiekoissa, meren pohjassa, syvältä louhitussa öljyssä ja jopa kuumissa lähteissä vettä, jonka lämpötila on noin 80 ºС. Ne elävät kasveilla, hedelmillä, erilaisilla eläimillä ja ihmisillä suolistossa, suussa, raajoissa ja kehon pinnalla.
Bakteerit ovat pienimpiä ja lukuisimpia eläviä olentoja. Pienen kokonsa ansiosta ne tunkeutuvat helposti halkeamiin, rakoihin, huokosiin. Erittäin kestävä ja sopeutunut erilaisiin olemassaolon olosuhteisiin. Ne sietävät kuivumista, äärimmäistä kylmyyttä, kuumenemista 90 ºС:een menettämättä elinkelpoisuutta.
Maapallolla ei käytännössä ole paikkaa, jossa bakteereita ei löytyisi, mutta eri määriä. Bakteerien elinolosuhteet ovat vaihtelevat. Jotkut heistä tarvitsevat ilman happea, toiset eivät sitä ja pystyvät elämään hapettomassa ympäristössä.
Ilmassa: bakteerit nousevat yläilmakehään jopa 30 km:n päähän. ja enemmän.
Varsinkin paljon niitä maaperässä. Yksi gramma maaperää voi sisältää satoja miljoonia bakteereja.
Vedessä: avoimien säiliöiden pintavesikerroksissa. Hyödylliset vesibakteerit mineralisoivat orgaanisia jäämiä.
Elävissä organismeissa: patogeeniset bakteerit pääsevät kehoon ulkoisesta ympäristöstä, mutta vain suotuisissa olosuhteissa aiheuttavat sairauksia. Symbioottiset elävät ruoansulatuselimissä, auttavat hajottamaan ja omaksumaan ruokaa, syntetisoivat vitamiineja.
Ulkoinen rakenne
Bakteerisolu on puettu erityiseen tiheään kuoreen - soluseinämään, joka suorittaa suojaavia ja tukitoimintoja ja antaa bakteerille myös pysyvän, tyypillisen muodon. Bakteerin soluseinä muistuttaa kasvisolun kuorta. Se on läpäisevä: sen kautta ravinteet kulkevat vapaasti soluun ja aineenvaihduntatuotteet lähtevät ympäristöön. Bakteerit kehittävät usein ylimääräisen suojaavan limakerroksen, kapselin, soluseinän päälle. Kapselin paksuus voi olla monta kertaa suurempi kuin itse kennon halkaisija, mutta se voi olla hyvin pieni. Kapseli ei ole pakollinen osa solua, se muodostuu riippuen olosuhteista, joissa bakteerit pääsevät sisään. Se estää bakteereja kuivumasta.
Joidenkin bakteerien pinnalla on pitkiä siimoja (yksi, kaksi tai monta) tai lyhyitä ohuita villoja. Siipien pituus voi olla monta kertaa suurempi kuin bakteerin kehon koko. Bakteerit liikkuvat lippujen ja villien avulla.

Sisäinen rakenne
Bakteerisolun sisällä on tiheä liikkumaton sytoplasma. Sillä on kerrosrakenne, tyhjiä ei ole, joten erilaiset proteiinit (entsyymit) ja vararavinteet sijaitsevat sytoplasman aineessa. Bakteerisoluissa ei ole ydintä. Niiden solujen keskiosaan on keskittynyt perinnöllistä tietoa kantava aine. Bakteerit, - nukleiinihappo - DNA. Mutta tämä aine ei ole kehystetty ytimeen.
Bakteerisolun sisäinen organisaatio on monimutkainen ja sillä on omat erityispiirteensä. Sytoplasma on erotettu soluseinästä sytoplasmisella kalvolla. Sytoplasmassa erotetaan pääaine eli matriisi, ribosomit ja pieni määrä kalvorakenteita, jotka suorittavat erilaisia toimintoja (mitokondrioiden analogit, endoplasminen retikulumi, Golgi-laite). Bakteerisolujen sytoplasmassa on usein erimuotoisia ja -kokoisia rakeita. Rakeet voivat koostua yhdisteistä, jotka toimivat energian ja hiilen lähteenä. Bakteerisoluista löytyy myös rasvapisaroita.
Solun keskiosassa ydinaine, DNA, on paikantunut, eikä sitä ole erotettu sytoplasmasta kalvolla. Tämä on ytimen analogi - nukleoidi. Nukleoidilla ei ole kalvoa, nukleolia eikä joukkoa kromosomeja.
Ravitsemusmenetelmät
Bakteereilla on erilaisia ruokintatapoja. Niiden joukossa ovat autotrofit ja heterotrofit. Autotrofit ovat organismeja, jotka voivat itsenäisesti muodostaa orgaanisia aineita ravintoonsa.
Kasvit tarvitsevat typpeä, mutta ne eivät itse pysty imemään typpeä ilmasta. Jotkut bakteerit yhdistävät ilmassa olevia typpimolekyylejä muihin molekyyleihin, jolloin saadaan kasveille saatavilla olevia aineita.
Nämä bakteerit asettuvat nuorten juurien soluihin, mikä johtaa juurien paksuuntumien muodostumiseen, joita kutsutaan kyhmyiksi. Tällaiset kyhmyt muodostuvat palkokasvien ja joidenkin muiden kasvien juuriin.

Juuret antavat bakteereille hiilihydraatteja ja bakteerit antavat juurille typpeä sisältäviä aineita, jotka kasvi voi ottaa itseensä. Heidän suhteensa hyödyttää molempia.
Kasvien juuret erittävät monia orgaanisia aineita (sokereita, aminohappoja ja muita), joita bakteerit ruokkivat. Siksi erityisesti monet bakteerit asettuvat juuria ympäröivään maakerrokseen. Nämä bakteerit muuttavat kuolleet kasvitähteet kasvin käytettävissä oleviksi aineiksi. Tätä maakerrosta kutsutaan risosfääriksi.
On olemassa useita hypoteeseja kyhmybakteerien tunkeutumisesta juurikudoksiin:
- epidermaalisen ja kortikaalisen kudoksen vaurioitumisen kautta;
- juurikarvojen läpi;
- vain nuoren solukalvon läpi;
- pektinolyyttisiä entsyymejä tuottavien seurabakteerien vuoksi;
- johtuen B-indolietikkahapon synteesin stimulaatiosta tryptofaanista, jota on aina läsnä kasvien juurieritteissä.
Kyhmybakteerien viemisprosessi juurikudokseen koostuu kahdesta vaiheesta:
- juurikarvojen infektio;
- kyhmyjen muodostumisprosessi.
Useimmissa tapauksissa tunkeutuva solu lisääntyy aktiivisesti, muodostaa niin sanottuja infektiolankoja ja siirtyy jo tällaisten säikeiden muodossa kasvikudoksiin. Infektiolangasta nousseet kyhmybakteerit jatkavat lisääntymistä isäntäkudoksessa.

Kasvien solut alkavat jakautua intensiivisesti, kun ne ovat täynnä nopeasti lisääntyviä kyhmybakteerisoluja. Nuoren kyhmyn yhdistäminen palkokasvin juureen tapahtuu vaskulaaristen kuitukimppujen ansiosta. Toiminnan aikana kyhmyt ovat yleensä tiheitä. Optimaalisen aktiivisuuden ilmenemiseen mennessä kyhmyt saavat vaaleanpunaisen värin (legoglobiinipigmentin vuoksi). Vain legoglobiinia sisältävät bakteerit pystyvät sitomaan typpeä.
Kyhmybakteerit tuottavat kymmeniä ja satoja kiloja typpilannoitteita hehtaaria kohden.
Aineenvaihdunta
Bakteerit eroavat toisistaan aineenvaihdunnassa. Joillekin se menee hapen mukana, toisille - ilman sen osallistumista.
Useimmat bakteerit ruokkivat valmiita orgaanisia aineita. Vain harvat niistä (sinivihreät tai syanobakteerit) pystyvät luomaan orgaanisia aineita epäorgaanisista. Niillä oli tärkeä rooli hapen kertymisessä maapallon ilmakehään.
Bakteerit imevät aineita ulkopuolelta, repivät molekyylinsä erilleen, kokoavat kuorensa näistä osista ja täydentävät niiden sisältöä (näin ne kasvavat) ja heittävät tarpeettomia molekyylejä ulos. Bakteerin kuori ja kalvo mahdollistavat sen, että se imee vain oikeat aineet.

Jos bakteerin kuori ja kalvo olisivat täysin läpäisemättömiä, mitään aineita ei pääsisi soluun. Jos ne olisivat läpäiseviä kaikille aineille, solun sisältö sekoittuisi väliaineeseen - liuokseen, jossa bakteeri elää. Bakteerien selviytymistä varten tarvitaan kuori, joka päästää tarvittavat aineet läpi, mutta ei niitä, joita ei tarvita.
Bakteeri imee ravinteita, jotka ovat lähellä sitä. Mitä tapahtuu seuraavaksi? Jos se voi liikkua itsenäisesti (siirrettä liikuttamalla tai työntämällä limaa takaisin), se liikkuu, kunnes se löytää tarvittavat aineet.

Jos se ei voi liikkua, se odottaa, kunnes diffuusio (yhden aineen molekyylien kyky tunkeutua toisen aineen molekyylien paksuun osaan) tuo siihen tarvittavat molekyylit.
Bakteerit yhdessä muiden mikro-organismiryhmien kanssa suorittavat valtavan kemiallisen työn. Muuntamalla erilaisia yhdisteitä ne saavat elintärkeää toimintaansa varten tarvittavan energian ja ravintoaineet. Aineenvaihduntaprosessit, energian saantitavat ja materiaalien tarve kehonsa aineiden rakentamiseksi bakteereihin ovat erilaisia.
Muut bakteerit tyydyttävät kaikki kehon orgaanisten aineiden synteesiin tarvittavat hiilen tarpeet epäorgaanisten yhdisteiden kustannuksella. Niitä kutsutaan autotrofeiksi. Autotrofiset bakteerit pystyvät syntetisoimaan orgaanisia aineita epäorgaanisista. Niistä erotetaan:
Kemosynteesi
Säteilyenergian käyttö on tärkein, mutta ei ainoa tapa luoda orgaanista ainetta hiilidioksidista ja vedestä. Tiedetään bakteereja, jotka eivät käytä auringonvaloa energialähteenä tällaiseen synteesiin, vaan organismien soluissa esiintyvien kemiallisten sidosten energiaa tiettyjen epäorgaanisten yhdisteiden - rikkivedyn, rikin, ammoniakin, vedyn, typpihapon, rautapitoisten yhdisteiden - hapettumisen aikana. rautaa ja mangaania. He käyttävät tätä kemiallista energiaa käyttämällä muodostunutta orgaanista ainetta kehonsa solujen rakentamiseen. Siksi tätä prosessia kutsutaan kemosynteesiksi.
Kemosynteettisten mikro-organismien tärkein ryhmä ovat nitrifioivat bakteerit. Nämä bakteerit elävät maaperässä ja suorittavat orgaanisten jäämien hajoamisen aikana muodostuneen ammoniakin hapettumisen typpihapoksi. Jälkimmäinen, reagoi maaperän mineraaliyhdisteiden kanssa, muuttuu typpihapon suoloiksi. Tämä prosessi tapahtuu kahdessa vaiheessa.
Rautabakteerit muuttavat rautaraudan oksidiksi. Muodostunut rautahydroksidi laskeutuu ja muodostaa niin sanotun suon rautamalmin.
Jotkut mikro-organismit ovat olemassa molekyylivedyn hapettumisen vuoksi, mikä tarjoaa autotrofisen ravintotavan.
Vetybakteereille tyypillinen piirre on kyky siirtyä heterotrofiseen elämäntapaan orgaanisten yhdisteiden kanssa ja vedyn puuttuessa.
Siten kemoautotrofit ovat tyypillisiä autotrofeja, koska ne syntetisoivat itsenäisesti tarvittavia orgaanisia yhdisteitä epäorgaanisista aineista eivätkä ota niitä valmiina muista organismeista, kuten heterotrofeista. Kemoautotrofiset bakteerit eroavat fototrofisista kasveista siinä, että ne ovat täysin riippumattomia valosta energianlähteenä.
bakteerien fotosynteesi
Jotkut pigmenttipitoiset rikkibakteerit (violetti, vihreä), jotka sisältävät tiettyjä pigmenttejä - bakterioklorofyllejä, pystyvät absorboimaan aurinkoenergiaa, jonka avulla rikkivety hajoaa organismeissaan ja antaa vetyatomeja vastaavien yhdisteiden palauttamiseksi. Tällä prosessilla on paljon yhteistä fotosynteesin kanssa, ja se eroaa vain siinä, että purppuraisissa ja vihreissä bakteereissa rikkivety (joskus karboksyylihapot) on vedyn luovuttaja ja vihreissä kasveissa vesi. Niissä ja muissa vedyn halkeaminen ja siirto tapahtuu absorboituneiden auringonsäteiden energian vuoksi.
Tällaista bakteerien fotosynteesiä, joka tapahtuu ilman hapen vapautumista, kutsutaan valopelkistykseksi. Hiilidioksidin valopelkistys liittyy vedyn siirtoon ei vedestä, vaan rikkivedystä:
6CO 2 + 12H 2 S + hv → C6H 12 O 6 + 12S \u003d 6H 2 O
Kemosynteesin ja bakteerien fotosynteesin biologinen merkitys planeetan mittakaavassa on suhteellisen pieni. Vain kemosynteettisillä bakteereilla on merkittävä rooli luonnon rikkikierrossa. Vihreiden kasvien imeytyminen rikkihapon suolojen muodossa, rikki palautuu ja siitä tulee osa proteiinimolekyylejä. Lisäksi kuolleiden kasvi- ja eläintähteiden tuhoamisen aikana mädäntäbakteerien toimesta rikki vapautuu rikkivedyn muodossa, jonka rikkibakteerit hapettavat vapaaksi rikiksi (tai rikkihapoksi), joka muodostaa kasvien käytettävissä olevia sulfiitteja maaperään. Kemo- ja fotoautotrofiset bakteerit ovat välttämättömiä typen ja rikin kierrossa.
itiöintiä
Itiöt muodostuvat bakteerisolun sisällä. Itiöiden muodostumisprosessissa bakteerisolu käy läpi sarjan biokemiallisia prosesseja. Vapaan veden määrä siinä vähenee, entsymaattinen aktiivisuus vähenee. Tämä varmistaa itiöiden kestävyyden epäsuotuisia ympäristöolosuhteita vastaan (korkea lämpötila, korkea suolapitoisuus, kuivuminen jne.). Itiöiden muodostuminen on ominaista vain pienelle bakteeriryhmälle.

Itiöt eivät ole olennainen vaihe bakteerien elinkaaressa. Itiöityminen alkaa vasta ravinteiden puutteesta tai aineenvaihduntatuotteiden kertymisestä. Itiöiden muodossa olevat bakteerit voivat pysyä lepotilassa pitkään. Bakteeri-itiöt kestävät pitkäaikaista keittämistä ja erittäin pitkäaikaista jäätymistä. Kun suotuisat olosuhteet syntyvät, riita itää ja tulee elinkelpoiseksi. Bakteeri-itiöt ovat mukautuksia selviytyäkseen epäsuotuisissa olosuhteissa.
jäljentäminen
Bakteerit lisääntyvät jakamalla yksi solu kahdeksi. Saavutettuaan tietyn koon bakteeri jakautuu kahteen identtiseen bakteeriin. Sitten jokainen heistä alkaa ruokkia, kasvaa, jakautua ja niin edelleen.

Solun pidentymisen jälkeen muodostuu vähitellen poikittainen väliseinä, ja sitten tytärsolut eroavat toisistaan; monissa bakteereissa tietyissä olosuhteissa solut pysyvät jakautumisen jälkeen kytkettyinä tunnusomaisiin ryhmiin. Tällöin syntyy erilaisia muotoja jakotason suunnasta ja jakojen lukumäärästä riippuen. Lisääntymistä silmuttamalla tapahtuu poikkeuksena bakteereissa.
Suotuisissa olosuhteissa solun jakautuminen tapahtuu monissa bakteereissa 20-30 minuutin välein. Tällaisella nopealla lisääntymisellä yhden bakteerin jälkeläiset voivat 5 päivässä muodostaa massan, joka voi täyttää kaikki meret ja valtameret. Yksinkertainen laskelma osoittaa, että 72 sukupolvea (720 000 000 000 000 000 000 solua) voidaan muodostaa päivässä. Jos käännetään painoksi - 4720 tonnia. Tätä ei kuitenkaan tapahdu luonnossa, koska useimmat bakteerit kuolevat nopeasti auringonvalon, kuivumisen, ruuan puutteen, 65-100 ºС:n lämpötilan vaikutuksesta, lajien välisen taistelun jne. seurauksena.
Bakteeri (1), joka on imenyt riittävästi ruokaa, kasvaa kokoonsa (2) ja alkaa valmistautua lisääntymiseen (solun jakautumiseen). Sen DNA (bakteerissa DNA-molekyyli on suljettu renkaaseen) kaksinkertaistuu (bakteeri tuottaa kopion tästä molekyylistä). Molemmat DNA-molekyylit (3.4) näyttävät kiinnittyneen bakteerin seinämään ja pidennettynä bakteerit hajaantuvat sivuille (5.6). Ensin nukleotidi jakautuu, sitten sytoplasma.
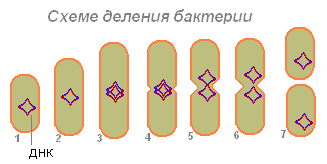
Kahden DNA-molekyylin hajoamisen jälkeen bakteereissa ilmaantuu supistelu, joka jakaa vähitellen bakteerin ruumiin kahteen osaan, joista jokainen sisältää DNA-molekyylin (7).
Se tapahtuu (heinäbasillissa), kaksi bakteeria tarttuu yhteen ja niiden välille muodostuu silta (1,2).

DNA kuljetetaan bakteerista toiseen hyppääjän (3) kautta. Yhdessä bakteerissa DNA-molekyylit kietoutuvat yhteen, tarttuvat yhteen paikoin (4), minkä jälkeen ne vaihtavat osia (5).
Bakteerien rooli luonnossa
Levikki
Bakteerit ovat tärkein lenkki yleisessä ainekierrossa luonnossa. Kasvit luovat monimutkaisia orgaanisia aineita hiilidioksidista, vedestä ja maaperän mineraalisuoloista. Nämä aineet palaavat maaperään kuolleiden sienten, kasvien ja eläinten ruumiiden mukana. Bakteerit hajottavat monimutkaisia aineita yksinkertaisiksi aineiksi, joita kasvit käyttävät uudelleen.
Bakteerit tuhoavat kuolleiden kasvien ja eläinten ruumiiden monimutkaisen orgaanisen aineen, elävien organismien eritteet ja erilaiset jätteet. Näillä orgaanisilla aineilla ruokkivat saprofyyttiset hajoamisbakteerit muuttavat ne humukseksi. Tällaisia ovat planeettamme järjestykset. Siten bakteerit osallistuvat aktiivisesti luonnon aineiden kiertoon.
maaperän muodostumista
Koska bakteereja on levinnyt lähes kaikkialle ja niitä löytyy valtavia määriä, ne määräävät suurelta osin erilaisia luonnossa tapahtuvia prosesseja. Syksyllä puiden ja pensaiden lehdet putoavat, maanpäälliset ruohon versot kuolevat, vanhat oksat putoavat ja aika ajoin vanhojen puiden rungot putoavat. Kaikki tämä muuttuu vähitellen humukseksi. 1 cm 3:ssa. Metsämaan pintakerros sisältää satoja miljoonia saprofyyttisiä maaperän bakteereja useista lajeista. Nämä bakteerit muuttavat humuksen erilaisiksi mineraaleiksi, jotka kasvien juuret voivat imeytyä maaperästä.
Jotkut maaperän bakteerit pystyvät imemään typpeä ilmasta hyödyntäen sitä elämänprosesseissa. Nämä typpeä sitovat bakteerit elävät yksinään tai asettuvat palkokasvien juuriin. Nämä bakteerit tunkeutuessaan palkokasvien juuriin aiheuttavat juurisolujen kasvua ja kyhmyjen muodostumista niihin.
Nämä bakteerit vapauttavat typpiyhdisteitä, joita kasvit käyttävät. Bakteerit saavat kasveista hiilihydraatteja ja kivennäissuoloja. Siten palkokasvien ja kyhmybakteerien välillä on läheinen suhde, joka on hyödyllinen sekä yhdelle että toiselle organismille. Tätä ilmiötä kutsutaan symbioosiksi.
Symbioosinsa kyhmybakteerien kanssa palkokasvit rikastavat maaperää typellä, mikä auttaa lisäämään satoa.
Jakautuminen luonnossa
Mikro-organismeja on kaikkialla. Ainoat poikkeukset ovat aktiivisten tulivuorten kraatterit ja pienet alueet räjähtäneiden atomipommien keskuksissa. Etelämantereen alhaiset lämpötilat, geysirien kiehuvat suihkut, suolaaltaiden kyllästetyt suolaliuokset, vuorenhuippujen voimakas insolaatio tai ydinreaktorien ankara säteily eivät häiritse mikroflooran olemassaoloa ja kehitystä. Kaikki elävät olennot ovat jatkuvasti vuorovaikutuksessa mikro-organismien kanssa, jotka eivät usein ole vain niiden varastoja, vaan myös jakelijoita. Mikro-organismit ovat planeettamme kotoisin, ja ne kehittävät aktiivisesti uskomattomimpia luonnollisia substraatteja.
Maaperän mikrofloora
Bakteerien määrä maaperässä on erittäin suuri - satoja miljoonia ja miljardeja yksilöitä 1 grammassa. Niitä on paljon enemmän maaperässä kuin vedessä ja ilmassa. Bakteerien kokonaismäärä maaperässä vaihtelee. Bakteerien määrä riippuu maaperän tyypistä, niiden kunnosta, kerrosten syvyydestä.
Maaperähiukkasten pinnalla mikro-organismit sijaitsevat pienissä mikropesäkkeissä (kukin 20-100 solua). Usein ne kehittyvät orgaanisen aineen hyytymien paksuuksissa, elävien ja kuolevien kasvien juurissa, ohuissa kapillaareissa ja kokkareiden sisällä.
Maaperän mikrofloora on hyvin monipuolinen. Täältä löytyy erilaisia fysiologisia bakteeriryhmiä: mätänevät, nitrifioivat, typpeä sitovat, rikkibakteerit jne. Niiden joukossa on aerobeja ja anaerobeja, itiö- ja ei-itiömuotoja. Mikrofloora on yksi maaperän muodostumisen tekijöistä.
Mikro-organismien kehitysalue maaperässä on elävien kasvien juurien vieressä oleva vyöhyke. Sitä kutsutaan risosfääriksi, ja sen sisältämien mikro-organismien kokonaisuutta kutsutaan risosfäärin mikroflooraksi.
Säiliöiden mikrofloora
Vesi on luonnollinen ympäristö, jossa mikro-organismeja kasvaa suuria määriä. Suurin osa niistä tulee veteen maaperästä. Tekijä, joka määrittää bakteerien määrän vedessä, ravinteiden läsnäolon siinä. Puhtaimmat ovat arteesisten kaivojen ja lähteiden vedet. Avoimet altaat ja joet ovat erittäin runsaasti bakteereja. Eniten bakteereja löytyy veden pintakerroksista, lähempänä rantaa. Kun etäisyys rannikosta kasvaa ja syvyys kasvaa, bakteerien määrä vähenee.
Puhdas vesi sisältää 100-200 bakteeria 1 ml:ssa, kun taas saastunut vesi sisältää 100-300 tuhatta tai enemmän. Pohjalieteessä on paljon bakteereja, erityisesti pintakerroksessa, jossa bakteerit muodostavat kalvon. Tässä kalvossa on paljon rikki- ja rautabakteereja, jotka hapettavat rikkivedyn rikkihapoksi ja estävät siten kalojen kuoleman. Lietessä on enemmän itiöitä kantavia muotoja, kun taas vedessä vallitsee itiöttömät muodot.
Lajikoostumukseltaan veden mikrofloora on samanlainen kuin maaperän mikrofloora, mutta myös erityisiä muotoja löytyy. Tuhoamalla erilaisia veteen pudonneita jätteitä mikro-organismit suorittavat vähitellen niin sanotun biologisen veden puhdistuksen.
Ilman mikrofloora
Ilman mikroflooraa on vähemmän kuin maaperän ja veden mikroflooraa. Bakteerit nousevat ilmaan pölyn mukana, voivat viipyä siellä jonkin aikaa ja asettua sitten maan pinnalle ja kuolevat ravinnon puutteesta tai ultraviolettisäteiden vaikutuksesta. Mikro-organismien määrä ilmassa riippuu maantieteellisestä alueesta, sijainnista, vuodenajasta, pölysaastuksesta jne. Jokainen pölyhiukkanen on mikro-organismien kantaja. Suurin osa bakteereista ilmassa teollisuusyritysten päällä. Maaseudulla ilma on puhtaampaa. Puhtain ilma on metsien, vuorten ja lumisten alueiden yllä. Ilman ylemmät kerrokset sisältävät vähemmän bakteereita. Ilman mikrofloorassa on monia pigmentoituneita ja itiöitä kantavia bakteereita, jotka kestävät muita paremmin ultraviolettisäteitä.
Ihmiskehon mikrofloora
Ihmisen keho, jopa täysin terve, on aina mikroflooran kantaja. Kun ihmiskeho joutuu kosketuksiin ilman ja maaperän kanssa, vaatteille ja iholle asettuu erilaisia mikro-organismeja, mukaan lukien taudinaiheuttajat (jäykkäkouristusbasillit, kaasukuolio jne.). Ihmisen altistuneet osat ovat useimmiten saastuneita. E. coli, stafylokokit löytyvät käsistä. Suuontelossa on yli 100 erilaista mikrobeja. Suun lämpötila, kosteus ja ravintojäämät ovat erinomainen ympäristö mikro-organismien kehittymiselle.
Vatsassa on hapan reaktio, joten suurin osa siinä olevista mikro-organismeista kuolee. Ohutsuolesta alkaen reaktio muuttuu emäksiseksi, ts. suotuisa mikrobeille. Paksusuolen mikrofloora on hyvin monipuolinen. Jokainen aikuinen erittää päivittäin noin 18 miljardia bakteeria ulosteiden mukana, ts. enemmän yksilöitä kuin ihmisiä maapallolla.
Sisäelimet, jotka eivät ole yhteydessä ulkoiseen ympäristöön (aivot, sydän, maksa, virtsarakko jne.), ovat yleensä vapaita mikrobeista. Mikrobit pääsevät näihin elimiin vain sairauden aikana.
Bakteerit pyöräilyssä
Mikro-organismeilla yleensä ja erityisesti bakteereilla on tärkeä rooli maapallon biologisesti tärkeissä aineiden kiertokuluissa, ja ne suorittavat kemiallisia muutoksia, jotka eivät ole täysin kasvien tai eläinten ulottumattomissa. Erityyppiset organismit suorittavat alkuaineiden kierron eri vaiheita. Kunkin erillisen organismiryhmän olemassaolo riippuu muiden ryhmien suorittamasta alkuaineiden kemiallisesta muutoksesta.
typen kierto
Typpiyhdisteiden syklisellä muuttumisella on ensiarvoisen tärkeä rooli tarvittavien typen muotojen toimittamisessa erilaisille biosfäärin organismeille ravitsemuksellisten tarpeiden kannalta. Yli 90 % typen kokonaissitoutumisesta johtuu tiettyjen bakteerien metabolisesta aktiivisuudesta.
Hiilen kiertokulku
Orgaanisen hiilen biologinen muuttuminen hiilidioksidiksi, johon liittyy molekyylihapen pelkistyminen, vaatii erilaisten mikro-organismien yhteistä metabolista toimintaa. Monet aerobiset bakteerit suorittavat orgaanisten aineiden täydellisen hapettumisen. Aerobisissa olosuhteissa orgaaniset yhdisteet hajotetaan aluksi käymisen avulla ja orgaanisen käymisen lopputuotteet hapetetaan edelleen anaerobisella hengityksellä, jos läsnä on epäorgaanisia vedyn vastaanottajia (nitraattia, sulfaattia tai CO2).
Rikkikierto
Eläville organismeille rikkiä on saatavana pääasiassa liukoisten sulfaattien tai pelkistettyjen orgaanisten rikkiyhdisteiden muodossa.
Raudan kierto
Jotkut makean veden säiliöt sisältävät suuria pitoisuuksia pelkistettyjä rautasuoloja. Tällaisissa paikoissa kehittyy erityinen bakteerimikrofloora - rautabakteerit, jotka hapettavat pelkistynyttä rautaa. Ne osallistuvat suon rautamalmien ja vesilähteiden muodostumiseen, joissa on runsaasti rautasuoloja.
Bakteerit ovat vanhimpia organismeja, ja ne ilmestyivät noin 3,5 miljardia vuotta sitten arkeaan. Noin 2,5 miljardia vuotta ne hallitsivat maapalloa muodostaen biosfäärin ja osallistuivat happiilmakehän muodostumiseen.
Bakteerit ovat yksi yksinkertaisimmin järjestetyistä elävistä organismeista (viruksia lukuun ottamatta). Niiden uskotaan olevan ensimmäisiä organismeja, jotka ilmestyivät maan päälle.
bakteerit- Nämä ovat yksisoluisia, klorofyllittomia organismeja, jotka lisääntyvät vegetatiivisesti jakautumalla, harvemmin nauhoittamalla, joskus muodostaen solunsisäisiä itiöitä.
Bakteerien koko mitataan mikroneina ja harvoja poikkeuksia lukuun ottamatta vaihtelee välillä 0,06-0,3 - 3-5 μ. Vesipisarassa voi helposti olla useita satoja miljoonia bakteereita.
Bakteerisolun muoto on melko tasainen. Bakteerien kolme päämuotoa tunnetaan: pyöreät, sauvan muotoiset ja kierteiset, joissa on lukuisia ja huomaamattomia siirtymiä niiden välillä. Anton De Bari vertasi niitä kuvaannollisesti biljardipalloon, kynään ja korkkiruuviin.
Bakteereja, jotka ovat muodoltaan pyöreitä, kutsutaan kokkeiksi. Ne eroavat koosta ja suhteellisesta sijainnista. Pareittain yhdistettyjä kokkeja kutsutaan diplokokiksi, mutta kaulakorun muodossa toisiinsa liittyneitä streptokokkeja. Jakaessaan, vuorotellen kahdessa keskenään kohtisuorassa suunnassa, muodostuu tetrakokkeja. Jos jako on oikea ja toistuu kolmessa keskenään kohtisuorassa suunnassa, soluyhteydet muodostuvat pakettien muodossa - nämä ovat niin sanottuja sardiineja. Jakaantuessaan eri suuntiin ilman suurta säännöllisyyttä, kokit muodostavat satunnaisia rypäleen sivellintä muistuttavia rypäleitä. Niitä kutsutaan stafylokokiksi.
Sauvan muotoiset bakteerit ovat ulkonäöltään hieman monipuolisempia. Ne voivat olla katkaistuja tai pyöristettyjä päitä, lieriömäisiä, tynnyrin muotoisia tai sitruunan muotoisia ja ikään kuin supisteita keskellä, usein ellipsoidisia, ja ne eroavat vain leveydeltä ja pituudeltaan. Tikut voivat olla suoria, kaarevia, yksittäisiä, pareittain tai ketjutettuja, lyhyitä tai voimakkaasti pitkänomaisia. Sauvan muotoisia bakteereja, joiden pituus on kaksi kertaa tai enemmän kuin leveys, kutsutaan basilleiksi; jos pituuden ja leveyden ero on pieni, niitä kutsutaan bakteereiksi.
Kiharamuotoiset bakteerit eroavat paitsi pituuden ja paksuuden lisäksi myös kiharoiden lukumäärästä ja luonteesta. Hieman kaarevia bakteereja (kihara ei ylitä 1/4 spiraalin kierrosta) kutsutaan vibrioiksi, ja bakteereja, joilla on yksi tai useampi suuri säännöllinen kihara, kutsutaan spirillumiksi. Pitkiä ja ohuita bakteereita, joilla on mutkainen muoto, jossa on lukuisia pieniä kiharoita, joskus koko filamentin suuria kaarevia, kutsutaan spirokeetiksi.
Bakteerien rakenne
Organisaation yksinkertaisuuden ja merkityksettömän kokonsa vuoksi bakteerit kuuluvat alkeellisimpiin olentoihin ja seisovat elämän alimmilla asteikoilla. Huolimatta tieteen ja tekniikan valtavasta edistymisestä, kaikkia bakteerien rakennetta koskevia kysymyksiä ei ole vielä ratkaistu.
Bakteerien runko koostuu eettisen sisällön kuoresta ja protoplasmasta, joka on kyllästetty solumehulla. Bakteerien kuori on ohut, väritön, sen rakennetta ei voida erottaa mikroskoopilla. Nähdäkseen sen he turvautuvat keinotekoisiin käsittelymenetelmiin. Suojus on solun ulkomuodon alla ja näyttää olevan tunnettu suoja haitallisia olosuhteita vastaan. Vapaasti päällystäen solun sisällön joustavuutensa ansiosta mahdollistaa bakteerien vapaan liikkeen, johon usein liittyy koko kehon reipas liikkeitä.
Vettä imevät kuoren ulommat kerrokset turpoavat usein ja muodostavat hyytelömäisen tahmean massan, joka saavuttaa huomattavan koon. Kun ulkokerrokset muuttuvat limaiseksi, kuori uusiutuu jatkuvasti protoplasman kustannuksella. Jäähdytettyä liimakuorta kutsutaan kapseliksi. Liman ja kapseleiden muodostumisen voimakkuus riippuu ravinnon ominaisuuksista ja voi joskus olla erittäin merkittävää. Limakapseli muodostuu paitsi jokaisen solun lähelle erikseen, myös moniin soluihin, jotka on yhdistetty yhdeksi pesäkkeeksi ja suljettu ikään kuin yhteiseen kapseliin. Näitä limaisia bakteeripesäkkeitä kutsutaan zoogleja. Kapseleiden muodostuminen ei ole ominaista kaikille bakteereille.
bakteerien liikkuminen
Kyky spontaaniin liikkumiseen on ominaista vain joillekin bakteeriryhmille. Bakteerit liikkuvat siimojen tai värekärojen avulla. Flagella näyttää enemmän tai vähemmän pitkiltä filamenteilta. Ne ovat erittäin herkkiä, ohuita, katkeavat helposti, eivätkä ne näy mikroskoopilla ilman erityistä värjäystä. Niiden halkaisija ei ylitä 1/20 bakteerirungon halkaisijasta.
Liikkuvat bakteerimuodot jaetaan seuraaviin ryhmiin:
- yksipuolinen - on vain yksi napainen siima,
- lophotrichous - solun toisessa päässä on nippu lippuja,
- peritrichous flagellat sijaitsevat koko kehon pinnalla.
Siipien sijainnin luonne bakteerin kehossa määrittää sen liikkeen luonteen - suoraviivainen tai satunnainen. Bakteerien liikkuvuus riippuu useista olosuhteista: lämpötilasta, ravintoalustan koostumuksesta, niiden elintärkeän toiminnan tuotteista jne. Liikkuvat bakteerimuodot eivät ole varustettu siimoilla kaikissa kehitysvaiheissaan eivätkä kaikissa kasvuolosuhteissa.
itiöintiä
Monien bakteerien kehossa esiintyy tiettyinä kehitysjaksoina pyöreitä tai ellipsoidisia muodostumia - tukia. Ne yleensä suorittavat bakteerien kehityssyklin loppuun. Itiöiden koko verrattuna niitä tuottaneiden solujen kokoon voi vaihdella suuresti.
Kaikentyyppisissä bakteereissa ei muodostu tukia. Niitä ympäröi hyvin eristetty kuori, joka on lähes vettä läpäisemätön ja ne ovat vakaimpia muodostelmia koko elävän maailman joukossa. Joten ne kestävät usein keittämistä useita tunteja ja pitkäaikaista altistumista kuivalle höyrylle 120-140 °:n lämpötiloissa. Monien basillien itiöt säilyttävät elinkelpoisuutensa sen jälkeen, kun ne ovat olleet pitkään -190 °:n lämpötilassa ja jopa nestemäisen vedyn lämpötilassa (-253 °). Ne kestävät myös kemikaalien - myrkkyjen - vaikutuksia. Kaikki tämä tekee erittäin vaikeaksi torjua patogeenisiä bakteeri-itiölajeja.
Kypsä itiö voi säilyä elinkelpoisena vuosikymmeniä. Yleensä itiöiden itäminen tapahtuu tietyn lepotilan jälkeen, ja se liittyy ulkoisten olosuhteiden vaikutukseen. Koko itiöitymisprosessi tapahtuu päivässä tai vähemmän. Itiöiden kypsymisen jälkeen niitä tuottanut solu kuolee vähitellen ja kypsät itiöt tulevat ulos. Itämisen aikana se turpoaa, rikastuu vedellä ja siitä tulee taimi pukeutunut ohueen kuoreen.
Bakteerien lisääntyminen
Saavutettuaan kypsyystilan ja kasvurajan bakteerit alkavat lisääntyä yksinkertaisella jakautumisella. Jakautumisen aikana bakteerin kehon keskiosaan ilmestyy väliseinä, joka sitten halkeaa ja erottaa kaksi uutta solua. Väliseinien järjestys jakautumisen aikana on erilainen eri bakteereille. Sauvan muotoisissa muodoissa se sijaitsee kohtisuorassa pitkää akselia vastaan, pallomaisissa muodoissa väliseinät voivat sijaita yhdessä, kahdessa tai kolmessa keskenään kohtisuorassa tasossa, mikä on syy sellaisten muotojen kuin streptokokkien, tetrakokkien ja sarkiinien muodostumiseen .
Bakteerien lisääntymisnopeus riippuu useista olosuhteista ja voi olla hyvinkin erilainen. Mitä suotuisammat ulkoiset olosuhteet bakteerien olemassaololle ovat, sitä nopeammin niiden jakautuminen tapahtuu. Normaaleissa olosuhteissa bakteerien määrä kaksinkertaistuu noin puolen tunnin välein. Jos se tapahtuisi aina esteettömästi, bakteerien määrä yhdestä solusta saavuttaisi valtavat mittasuhteet. Mikrobiologi Kohnin mukaan yhden basillin jälkeläinen viidessä päivässä voisi täyttää kaikki meret ja valtameret. Tällaista ei kuitenkaan ole koskaan tapahtunut eikä tule tapahtumaan. Bakteerien elinkaarta rajoittavat tietyt ulkoiset olosuhteet, joiden jälkeen lisääntyminen hidastuu tai pysähtyy kokonaan. Ravinnon puute, haitalliset aineenvaihduntatuotteet, eri lajien kilpailu jne. vaikuttavat haitallisesti bakteereihin. Epäsuotuisissa olosuhteissa ne kuolevat massiivisesti.
Bakteerien luokitus
Bakteerien asemaa elävien olentojen järjestelmässä ei ole vielä tarkasti määritelty. On yleisesti hyväksyttyä, että bakteerit ovat osa kasvimaailmaa, ja sienet ja levät ovat niitä lähimpiä sukulaisia organismeja. Bakteerien morfologiset ominaisuudet rajoittuvat useimmissa tapauksissa muutamiin muotoihin: pallomaiset, sauvat, spiraalit. Niiden ulkoisen organisaation poikkeuksellisen yksinkertaisuus ja alkeellisuus vaikeuttaa niiden luokittelua. Bakteerin lajin määrittäminen pelkästään morfologisten ominaisuuksien perusteella on mahdotonta. Tieteellinen systematiikka perustuu morfologiaan ja kehityshistoriaan, mutta bakteerien luokitteluun tarvitaan paitsi morfologia, myös niiden fysiologiset ja biokemialliset ominaisuudet. Tältä osin vahvistetaan seuraavat: bakteerien suhde happeen, lämpötilaolosuhteet, pigmentin muodostuminen, gelatiinin nesteyttäminen, happojen ja kaasun muodostuminen sokereihin, maidon muutos bakteerien kasvun aikana, indolin, rikkivedyn, ammoniakin muodostuminen, nitraattien pelkistäminen nitriiteiksi tai vapaaksi typeksi. Tämä ei kuitenkaan aina riitä bakteerityypin määrittämiseen.
Bakteereille on olemassa erilaisia luokitusjärjestelmiä, mutta ne ovat kaikki ehdollisia ja paljon enemmän tai vähemmän luonnollisesta luokittelusta poikkeavia. Näiden järjestelmien tai ainakin yhden niistä huomioon ottaminen ei ole tässä tapauksessa välttämätöntä edes fytopatogeenisiin bakteereihin sovellettaessa. On vain sanottava, että tällä hetkellä lähes kaikki fytopatogeeniset bakteerit yhdistyvät suvuiksi Pseudomonas, Xanthomonas, Bacterium ja Erwinia.
Äskettäin M. V. Gorlenko (1961) ehdotti seuraavaa luokitusjärjestelmää Eubacteriales-luokan fytopatogeenisille bakteereille:
minä Heimo Mycobacteriaceae(Chester, 1901) - liikkumattomat bakteerit (ilman siimoja):
- 1. suku - Gorynebacterium (Leman ja Neumann, 1896) - (gram-positiiviset bakteerit;
- 2. suvun Aplanobacterium (Smith, 1905, Geshich, 1956) - gramnegatiiviset bakteerit.
II. Heimo Pseudomonadaceae(Wilson et al., 1917) - bakteerit, joissa on flagella (polaarinen):
- 1. suku - Pseudomonas (Migula, 1900) - värjäämättömät ja fluoresoivat bakteerit;
- 2. suku - Xanthomonas (Dawson, 1839) - bakteerit, joissa on värillisiä pesäkkeitä.
III. Heimo Bakteerit(Kon, 1872) - liikkuvat bakteerit, joissa on peritrichous flagella, jotka eivät muodosta tukia:
- 1. suku - Bacterium (Ehrenberger, 1828) - värjäämättömät muodot, jotka eivät muodosta pektinaaseja ja protopektinaaseja;
- 2. suku - Pectobacterium (Waldy, 1945) - värjäämättömät muodot, jotka muodostavat pektinaasia ja protopektinaasia;
- 3. suku - Chromobacterium (Bergonzini, 1881) - värilliset muodot.
IV. Suku Bacillaceae(Fischer, 1895) - liikkuvat bakteerit, itiöitä muodostavat sauvat:
- 1. suku - Bacillus (Kon, 1832) - solut eivät turpoa tai turpoa hieman itiöiden muodostumisen aikana;
- 2. suku - Clostridium (Praznovsky, 1880) - solut turpoavat itiöiden muodostumisen aikana.
Yllä olevassa järjestelmässä tähän asti yleisesti hyväksytty Erwinia-suku on jätetty pois. Siitä eristetään erityinen Pectobacterium-suku, joka sisältää bakteereita, joilla on peritrichous flagella ja pectolyyttinen aktiivisuus. Ne fytopatogeeniset bakteerit, joilla ei ole tätä kykyä, luokitellaan Bacterium-sukuun. Tämä sinänsä rationaalinen järjestelmä on uusi eikä ole vielä astunut arkeen, joten noudatamme tässä työssä luokittelua, jossa Erwinia-suvulle on annettu paikka. Tätä kasvipatogeenisten bakteerien yleisnimeä käytetään laajalti erikoiskirjallisuudessa sekä maassamme että ulkomailla.
Bakteerityypin määrittäminen ilman keinotekoisia ravintoalustoja on mahdotonta. Tässä suhteessa voidaan todeta, että viljeltäessä bakteereja ne muodostavat erittäin tyypillisiä pesäkkeitä. Tässä tapauksessa yhtä ulkonäköä voidaan käyttää arvioimaan bakteerilajeja.
Jos löydät virheen, korosta tekstinpätkä ja napsauta Ctrl+Enter.
1.5. Nykyaikainen bakteeriluokitus Bakteerien modernissa taksonomiassa on kehittynyt tilanne, joka on tyypillinen myös muiden organismien luokittelulle: on edistytty sellaisen fylogeneettisen luokittelujärjestelmän luomisessa, joka heijastaa evolutionaarisen kehityksen pääsuuntia ja edustajien välistä suhdetta. tietyt taksonit, mutta keinotekoiset ilmiöt säilyttävät merkityksensä - tyypilliset luokitukset, kätevämpi mikro-organismien tunnistamisessa. Tällä hetkellä ei ole olemassa mitään yksityiskohtaista prokaryoottien evoluutiojärjestelmää, ja todennäköisesti tämän ongelman ratkaisu ei ole lähitulevaisuudessa. Prokaryoottien ominaisuudet morfologisen, fysiologisen, biokemiallisen ja geneettisen organisaation alalla osoittavat, että korkeampien organismien järjestelmän rakentamisessa käytettyjä hyvin kehittyneitä periaatteita ei voida soveltaa niihin. Käsittelemättä bakteerien taksonomian ongelman historiallisia näkökohtia, on huomattava, että hyväksyttävin fylogeneettinen luokittelujärjestelmä prokaryooteille on järjestelmä, joka perustuu 16S rRNA:n nukleotidisekvenssin yhteensovittamiseen. Tämä järjestelmä on pohjana moniosaisen prokaryoottien tietosanakirjan - Bergeyn käsikirjalle Systematic Bacteriology (Bergeyn opas systematic bakteereihin) 2. painokselle, jonka ensimmäinen osa julkaistiin vuonna 2001. Tässä työssä kaikki prokaryootit on jaettu 26 fylogeneettistä "haaraa" (ryhmää) perustuen niiden 16S-rRNA:n rakenteeseen; 23 "haaraa" edustavat eubakteerit ja kolmea arkkibakteerit. On korostettava, että suuri osa näistä fylogeneettisistä ryhmistä sisältää prokaryoottisia lajeja, joita ei ole eristetty puhdasviljelminä ja siksi niitä ei ole vielä tutkittu yksityiskohtaisesti. Näiden lajien edustajille tunnetaan tällä hetkellä vain 16S rRNA:n nukleotidisekvenssit. 23 eubakteeriryhmästä kahta fylogeneettistä ryhmää edustavat grampositiiviset bakteerit, loput ryhmät ovat gramnegatiivisia. Gram-negatiiviset bakteerit koostuvat suuresta ryhmästä proteobakteereja (Proteobacteria) ja 20 ryhmästä muita bakteereja, joilla on tämän tyyppinen soluseinä. Taulukossa 1 on lyhyt kuvaus proteobakteereista, joita useimpien eukaryoottien mitokondriot ja kloroplastit ovat lähimpänä 16S-rRNA-koostumuksessa. 2. Proteobakteerit ovat gram-negatiivisten bakteerien ryhmä, joka on hyvin heterogeeninen morfologisesti, fysiologisesti ja biokemiallisesti. Tämän ryhmän edustajille on ominaista kaikenlainen energia-aineenvaihdunta ja ravitsemus. Useimpien proteobakteerilajien solut ovat sauvan muotoisia, pallomaisia tai vibrioidisia, ja ne lisääntyvät pääasiassa binäärifissiolla, mutta joillekin lajeille on ominaista silmujen muodostuminen ja hedelmäkappaleiden muodostuminen monimutkaisessa solusyklissä. Tämä ryhmä sisältää sekä lippusolujen aiheuttamaa liikkuvuutta että liikkumattomia bakteereja. Molekyylihapen suhteen proteobakteerit luokitellaan pakollisiksi aerobeiksi, pakollisiksi ja fakultatiivisiksi anaerobeiksi. Proteobakteeriryhmä on jaettu viiteen alaryhmään 16S rRNA:n erojen perusteella: alfa, beeta, gamma, delta ja epsilon. Gram-negatiivisia ovat proteobakteerien lisäksi seuraavat eubakteerien pääryhmät: vetytermofiilit, vihreät rihmabakteerit, vihreät rikkibakteerit, syanobakteerit, spirokeetit, sytofaagit, bakteroidit, klamydia, planktomykeetit, deinokokki, chloroflexus, fusobakteerit, termobakteerit jne. Fylogeneettiset ryhmät grampositiiviset bakteerit - Actinobacteria ja Firmicutes. Actinobacteria-ryhmää ("actinomycete haara") edustavat seuraavat bakteerisuvut, joiden DNA:ssa on runsaasti GC-pareja: Geodermatophilus, Frankia, Streptomyces, Arthrobacter, Micrococcus, Actinomyces, Bifidobacterium, Propionibacterium, Actinoplanes, Nocardia, Cordococcus , Mycobacterium. Firmicutes-ryhmä ("klostridihaara" - pääasiassa grampositiiviset bakteerit, joiden GC-parien pitoisuus DNA:ssa on alhainen) koostuu seuraavista suvuista: Clostridium, Lactococcus, Pediococcus, Streptococcus, Enterococcus, Leuconostoc, Listeria, Caryophanon, Staphylococcus, Sarcinaccus , Sporosarcina, Bacillus, Desulfotomaculum, Heliobacterium, Mycoplasma, Ureaplasma jne. Arkkibakteerit jaetaan kolmeen fylogeneettiseen ryhmään: Crenarchaeota, Euryarchaeota ja Korarchaeota. Crenarchaeota-ryhmä koostuu erittäin termofiilisistä bakteereista, joiden edustajat useimmat metaboloivat rikkiä, jotkut vähentävät rauta- ja molybdeeni-ioneja. Euryarchaeota-ryhmään kuuluvat obligaatit anaerobiset metanogeeniset arkkibakteerit sekä äärimmäiset termofiilit ja halofiilit. Korarchaeota-ryhmän muodostavat kuumissa rikkilähteissä elävät arkebakteerit. Tähän mennessä ketään tämän ryhmän edustajia (joilla on samanlainen 16S-rRNA) ei ole eristetty puhdasviljelmänä, joten niiden fenotyyppisiä piirteitä ei ole tutkittu tarpeeksi. Prokaryoottien fylogeneettisten haarojen tarkastelun päätteeksi on huomattava, että ehdotettu fylogeneettinen järjestelmä, joka perustuu vain yhden ribosomaalisen RNA-geenin nukleotidisekvenssien tutkimukseen, ei ole muuta kuin yksi teknisesti kätevistä ja kehitetyistä järjestelmistä useiden tilausten järjestämiseen. organismeja niiden tunnistamiseksi, joten on loogista rakentaa oikea bakteeritaksonomia, kun tämä piirre ei ole mahdollista. Tunnetuin ja käytetyin bakteerien fenotyyppinen luokitus on Burgey's Guide to Bacteria -oppaan yhdeksännessä painoksessa esitetty. Tässä painoksessa bakteerit jaetaan neljään pääluokkaan (jakaumaan) solun rajakerroksen rakenteen perusteella: 1) Gracilicutes (latinasta cutes - skin, gracilis - ohut) - gramnegatiiviset eubakteerit, joilla on soluseinä; 2) Firmicutes (lat. firmus - vahva) - grampositiiviset eubakteerit, joilla on soluseinä; 3) Tenericutes (latinasta tener - pehmeä, herkkä) - eubakteerit, joissa ei ole soluseinämiä; 4) Mendosicutes (latinan sanasta mendosus - virheellinen) - arkkibakteerit, joiden soluseinät eroavat muiden prokaryoottien vastaavista rakenteista. Gracilicutes-osastolla on eri morfologioita omaavia bakteereja, joilla on gramnegatiivinen soluseinä. Lisääntyminen tapahtuu pääasiassa binäärifissiolla, jotkut bakteerit lisääntyvät orastumalla. Endospore ei muodostu. Suurin osa niistä on liikkuvia: bakteerit liikkuvat kaikentyyppisesti - lippujen avulla, liukumalla, taipumalla. Divisioonaan kuuluvat aerobiset, anaerobiset ja fakultatiiviset anaerobiset bakteerit; fototrofiset ja kemotrofiset bakteerit. Osasto on jaettu kolmeen luokkaan: Scotobacteria, Oxyphotobacteria, Anoxyphotobacteria. Luokkaan Scotobacteria kuuluvat gramnegatiiviset bakteerit, jotka eivät käytä valoenergiaa aineenvaihduntaan, vaan saavat sitä vain redox-reaktioiden seurauksena. Luokan nimi tulee kreikan kielestä scotos - pimeys. Tämä on suurin bakteeriluokka. Anoksifotobakteerien luokkaan kuuluvat purppurabakteerit, vihreät bakteerit ja heliobakteerit, jotka suorittavat happipitoista fotosynteesiä (ilman molekyylisen hapen vapautumista). Oksifotobakteerien luokkaa edustavat syanobakteerit ja proklorofyytit, jotka suorittavat happifotosynteesiä (vapauttamalla molekyylistä happea). Tämän tyyppinen fotosynteesi on samanlainen kuin kasveissa tapahtuva fotosynteesi. Firmicutes-osastolla on bakteereja, joilla on grampositiivinen soluseinä. Soluilla voi olla eri muotoja: sauvoja, kokkeja, rihmamaisia, haarautuvia. Jotkut edustajat muodostavat endosporeja. Useimmat heistä ovat liikkumattomia; liikkuvilla muodoilla on peritrichous flagella. Osastolla on aerobisia, anaerobisia ja fakultatiivisia anaerobisia bakteereja. Osastossa on kaksi luokkaa: Firmibakteerit, Tallobakteerit. Firmibakteeri-luokka sisältää suuren määrän "haarautumattomia" grampositiivisia bakteereja. Tallobakteerien luokkaan kuuluvat bakteerit, joiden solut pystyvät "haarautumaan". Tenericutes-osastoa edustavat bakteerit, joilla ei ole soluseinää. Soluseinän puuttumisen vuoksi solujen muoto ei ole vakio: yhden lajin puhdasviljelmässä esiintyy samanaikaisesti kokkoidi-, sauva-, rihma-, päärynä-, kiekko- ja muita soluja. Tähän osioon sisältyvien bakteerien lisääntyminen tapahtuu binäärifissiolla, orastumalla. Gramin tahra on negatiivinen. Tyypillistä on pienten, agariksi kasvavien pesäkkeiden muodostuminen. Ne voivat olla saprofyyttisiä, loisia tai patogeenisiä. Osasto koostuu yhdestä Mollicutes-luokasta (mykoplasmoista). Mendosicutes-jaon muodostavat bakteerit, joilla on jäykkä soluseinä, mutta joista puuttuu mureiinipeptidoglykaani. Useimmat edustajat ovat tiukkoja anaerobeja, joista monilla on flagella. Lajeille on ominaista ekologinen ja metabolinen monimuotoisuus, kyky elää äärimmäisissä olosuhteissa. Osasto koostuu yhdestä luokasta - Archaebacteria. Osana neljää jakoa (pääkategoriaa) on tunnistettu 35 bakteeriryhmää (tai jaksoa), joita kuvataan enemmän tai vähemmän seuraavissa luvuissa. Seuraavat ryhmät kuuluvat Gracilicutes-osastoon. Ryhmä 1. Spirochetes. Ryhmä 2. Aerobiset (tai mikroaerofiiliset), liikkuvat, spiraaliset (tai vibrioidiset) gramnegatiiviset bakteerit. Ryhmä 3. Liikkumattomat tai harvoin liikkuvat Gram-negatiiviset kaarevat bakteerit. Ryhmä 4. Gram-negatiiviset aerobiset (tai mikroaerofiiliset) sauvat ja kokit. Ryhmä 5. Valinnaiset aerobiset gram-negatiiviset sauvat. Ryhmä 6. Gram-negatiiviset anaerobiset suorat, kaarevat tai kierretangot. Ryhmä 7. Bakteerit, jotka suorittavat rikin tai sulfaatin dissimilaatiopelkistystä. Ryhmä 8. Anaerobiset gramnegatiiviset kokit. Ryhmä 9. Rickettsia ja klamydia. Ryhmä 10. Happittomat fototrofiset bakteerit. Ryhmä 11. Happipitoiset fototrofiset bakteerit. Ryhmä 12. Aerobiset kemolitotrofiset bakteerit ja niihin liittyvät organismit. Ryhmä 13. Orastelevat ja (tai) uloskasvua muodostavat bakteerit. Ryhmä 14. Bakteerit, joissa on vaipat. Ryhmä 15. Ei-fotosynteettiset liukubakteerit, jotka eivät muodosta hedelmäkappaleita. Ryhmä 16. Hedelmäkappaleita muodostavat liukubakteerit. Firmicutes-osastoon kuuluu: Ryhmä 17. Gram-positiiviset kokit. Ryhmä 18. Gram-positiiviset sauvat ja kokit, jotka muodostavat endosporeja. Ryhmä 19. Oikean muotoiset grampositiiviset sauvat, jotka eivät muodosta itiöitä. Ryhmä 20. Epäsäännöllisen muotoiset grampositiiviset sauvat, jotka eivät muodosta itiöitä. Ryhmä 21. Mykobakteerit. Ryhmät 22-29. Actinomycetes. Tenericutes-osastoon kuuluu: Ryhmä 30. Mykoplasmat. Division Mendosicutes sisältää: Ryhmä 31. Methanogens. Ryhmä 32. Sulfaattia vähentävät arkebakteerit. Ryhmä 33. Erittäin halofiiliset arkkibakteerit (halobakteerit). Ryhmä 34. Arkebakteerit, joista puuttuu soluseinä. Ryhmä 35 Erittäin termofiiliset ja hypertermofiiliset rikkiä metaboloivat arkebakteerit. Lopuksi on korostettava, että suurin osa luonnollisissa yhteisöissä esiintyvistä mikro-organismeista on vielä eristettävä puhdasviljelmiin. Uskotaan, että tällä hetkellä vain 0,1 % kokonaismikrobien monimuotoisuudesta voidaan viljellä, eikä ole mahdollista kasvattaa ja tunnistaa jäljellä olevia bakteerien edustajia, vaikka noin 5 tuhatta prokaryoottilajia on jo eristetty ja kuvattu puhtaassa viljelmässä.