Sammansättning av en DNA-nukleotid. Nukleotider. Förening. Strukturera. Sammansättning av nukleinsyror
Deoxiribonukleinsyror (DNA) är en makromolekyl (en av de tre huvudsakliga, de andra två är RNA och proteiner), som tillhandahåller lagring, överföring från generation till generation och implementering av det genetiska programmet för utveckling och funktion av levande organismer. DNA innehåller information om strukturen hos olika typer av RNA och proteiner.
I eukaryota celler (djur, växter och svampar) finns DNA i cellkärnan som en del av kromosomerna, liksom i vissa cellorganeller (mitokondrier och plastider). I cellerna hos prokaryota organismer (bakterier och archaea) är en cirkulär eller linjär DNA-molekyl, den så kallade nukleoiden, fäst från insidan till cellmembranet. De och lägre eukaryoter (till exempel jäst) har också små autonoma, mestadels cirkulära DNA-molekyler som kallas plasmider. Dessutom kan enkel- eller dubbelsträngade DNA-molekyler bilda genomet av DNA-innehållande virus.
Ur kemisk synvinkel är DNA en lång polymer molekyl som består av repeterande block - nukleotider. Varje nukleotid består av en kvävebas, ett socker (deoxiribos) och en fosfatgrupp. Bindningarna mellan nukleotider i en kedja bildas av deoxiribos och en fosfatgrupp (fosfodiesterbindningar). I den överväldigande majoriteten av fallen (förutom vissa virus som innehåller enkelsträngat DNA) består DNA-makromolekylen av två kedjor som är orienterade av kvävehaltiga baser till varandra. Denna dubbelsträngade molekyl är spiralformad. I allmänhet kallas DNA-molekylens struktur "dubbelhelixen".
Att dechiffrera DNA:s struktur (1953) var en av vändpunkterna i biologins historia. Francis Crick, James Watson och Maurice Wilkins tilldelades Nobelpriset i fysiologi eller medicin 1962 för sina enastående bidrag till denna upptäckt. Rosalind Franklin, som fick röntgenbilderna utan vilka Watson och Crick inte skulle ha kunnat dra slutsatser om DNA-strukturen, dog 1958 i cancer, och Nobelpriset ges inte postumt.
Ribonukleinsyror (RNA) är en av de tre huvudsakliga makromolekylerna (de andra två är DNA och proteiner) som finns i cellerna hos alla levande organismer.
Precis som DNA (deoxiribonukleinsyra) är RNA uppbyggt av en lång kedja där varje länk kallas en nukleotid. Varje nukleotid består av en kvävebas, ett ribossocker och en fosfatgrupp. Nukleotidernas sekvens tillåter RNA att koda för genetisk information. Alla cellulära organismer använder RNA (mRNA) för att programmera proteinsyntes.
Cellulärt RNA bildas under en process som kallas transkription, det vill säga syntesen av RNA på en DNA-mall, utförd av speciella enzymer - RNA-polymeraser. Messenger-RNA (mRNA) deltar sedan i en process som kallas translation. Translation är syntesen av ett protein på en mRNA-mall med deltagande av ribosomer. Andra RNA genomgår kemiska modifieringar efter transkription, och efter bildandet av sekundära och tertiära strukturer utför de funktioner som beror på typen av RNA.
Enkelsträngade RNA kännetecknas av en mängd olika rumsliga strukturer där några av nukleotiderna i samma kedja är parade med varandra. Vissa högstrukturerade RNA är involverade i cellproteinsyntes, till exempel tjänar överförings-RNA till att känna igen kodon och levererar motsvarande aminosyror till platsen för proteinsyntes, medan ribosomala RNA fungerar som den strukturella och katalytiska basen för ribosomer.
Funktionerna hos RNA i moderna celler är dock inte begränsade till deras roll i translation. Sålunda är små nukleära RNA involverade i splitsningen av eukaryota budbärar-RNA och andra processer.
Förutom det faktum att RNA-molekyler är en del av vissa enzymer (till exempel telomeras), har vissa RNA sin egen enzymatiska aktivitet: förmågan att göra brytningar i andra RNA-molekyler eller, omvänt, "limma" två RNA-fragment. Sådana RNA kallas ribozymer.
Genomen hos ett antal virus består av RNA, det vill säga i dem spelar det den roll som DNA spelar i högre organismer. Baserat på mångfalden av RNA-funktioner i cellen lades en hypotes fram, enligt vilken RNA är den första molekylen som var kapabel till självreproduktion i prebiologiska system.
Det finns tre huvudsakliga skillnader mellan DNA och RNA:
- 1. DNA innehåller sockret deoxiribos, RNA innehåller ribos, som har en extra hydroxylgrupp jämfört med deoxiribos. Denna grupp ökar sannolikheten för hydrolys av molekylen, det vill säga den minskar stabiliteten hos RNA-molekylen.
- 2. Nukleotiden som är komplementär till adenin i RNA är inte tymin, som i DNA, utan uracil är den ometylerade formen av tymin.
- 3. DNA finns i form av en dubbelhelix, bestående av två separata molekyler. RNA-molekyler är i genomsnitt mycket kortare och övervägande enkelsträngade.
Strukturanalys av biologiskt aktiva RNA-molekyler, inklusive tRNA, rRNA, snRNA och andra molekyler som inte kodar för proteiner, visade att de inte består av en lång helix, utan av många korta helixar som ligger nära varandra och bildar något liknande tertiär struktur av proteinet. Som ett resultat kan RNA katalysera kemiska reaktioner, till exempel består peptidyltransferascentrum i ribosomen, som är involverat i bildandet av peptidbindningen av proteiner, helt av RNA
År 1944 upptäckte O. Avery och hans kollegor K. McLeod och M. McCarthy den transformerande aktiviteten hos DNA i pneumokocker. Dessa författare fortsatte Griffiths arbete, som beskrev fenomenet transformation (överföring av ärftliga egenskaper) hos bakterier. O. Avery, K. McLeod, M. McCarthy visade att när proteiner, polysackarider och RNA avlägsnas störs inte omvandlingen av bakterier, och när den inducerande substansen exponeras för enzymet deoxiribonukleas försvinner den transformerande aktiviteten.
I dessa experiment visades DNA-molekylens genetiska roll för första gången. 1952 bekräftade A. Hershey och M. Chase DNA-molekylens genetiska roll i experiment på T2-bakteriofagen. Genom att markera dess protein med radioaktivt svavel och dess DNA med radioaktivt fosfor, infekterade de E. coli med detta bakteriella virus. I fagens avkomma hittades en stor mängd radioaktivt fosfor och endast spår av S. Det följde att det var DNA, och inte fagproteinet, som trängde in i bakterien och sedan, efter replikation, överfördes till fagavkomman. .
Strukturen av en DNA-nukleotid. Typer av nukleotider.
Nukleotid DNA består av
Kvävebas (4 typer i DNA: adenin, tymin, cytosin, guanin)
Monosocker deoxiribos
Fosforsyra
nukleotidmolekyl består av tre delar - ett socker med fem kolatomer, en kvävebas och fosforsyra.
Socker ingår i nukleotidsammansättning, innehåller fem kolatomer, det vill säga det är en pentos. Beroende på vilken typ av pentos som finns i nukleotiden, finns det två typer av nukleinsyror - ribonukleinsyror (RNA), som innehåller ribos, och deoxiribonukleinsyror (DNA), som innehåller deoxiribos. I deoxiribos ersätts OH-gruppen vid den andra kolatomen med en H-atom, det vill säga den har en syreatom mindre än i ribos.
I båda typer av nukleinsyror innehåller baser av fyra olika typer: två av dem tillhör klassen puriner och två till klassen pyrimidiner. Kvävet som ingår i ringen ger huvudkaraktären till dessa föreningar. Puriner inkluderar adenin (A) och guanin (G), och pyrimidiner inkluderar cytosin (C) och tymin (T) eller uracil (U) (i DNA respektive RNA). Tymin är kemiskt mycket nära uracil (det är 5-metyluracil, det vill säga uracil, där en metylgrupp finns vid den 5:e kolatomen). Purinmolekylen har två ringar, medan pyrimidinmolekylen har en.
Nukleotider är sammanlänkade genom en stark kovalent bindning genom sockret i en nukleotid och fosforsyran i en annan. Det visar sig polynukleotidkedja. I ena änden finns fri fosforsyra (5'-änden), i den andra är fritt socker (3'-änden). (DNA-polymeras kan bara lägga till nya nukleotider till 3'-änden.)
Två polynukleotidkedjor är förbundna med varandra genom svaga vätebindningar mellan kvävehaltiga baser. Det finns 2 regler:
komplementaritetsprincipen: tymin är alltid motsatsen till adenin, guanin är alltid motsatsen till cytosin (de matchar varandra i form och antal vätebindningar - det finns två bindningar mellan A och G, och 3 mellan C och G).
principen om antiparallelism: där en polynukleotidkedja har en 5'-ände, den andra har en 3'-ände och vice versa.
Det visar sig dubbel kedja DNA.
Hon vrider sig in i dubbel helix, ett varv av helixen har en längd av 3,4 nm, innehåller 10 par nukleotider. Kvävehaltiga baser (bevarare av genetisk information) finns inuti helixen, skyddade.
Nukleinsyror, liksom proteiner, är avgörande för livet. De representerar det genetiska materialet för alla levande organismer, ner till de enklaste virusen. Namnet "nukleinsyror" speglar det faktum att de är lokaliserade huvudsakligen i kärnan (kärna - kärna). Med specifik färgning för nukleinsyror är kärnorna mycket tydligt synliga i ett ljusmikroskop.
Ta reda på strukturen av DNA(deoxiribonukleinsyra) - en av de två befintliga typerna av nukleinsyror - öppnade en ny era inom biologin, eftersom den äntligen gjorde det möjligt att förstå hur levande organismer lagrar den information som behövs för att reglera deras liv och hur de överför denna information till sin avkomma . Vi har redan noterat ovan att nukleinsyror är sammansatta av monomera enheter som kallas nukleotider. Extremt långa molekyler - polynukleotider - är byggda av nukleotider.
För att förstå strukturen av polynukleotider är det därför nödvändigt att först bekanta sig med hur byggda nukleotider.
Nukleotider. Strukturen av nukleotider
nukleotidmolekyl består av tre delar - ett socker med fem kolatomer, en kvävebas och en fosforhaltig.
Socker ingår i nukleotidsammansättning, innehåller fem kolatomer, det vill säga det är en pentos. Beroende på vilken typ av pentos som finns i nukleotiden, finns det två typer av nukleinsyror - ribonukleinsyror (RNA), som innehåller ribos, och deoxiribonukleinsyror (DNA), som innehåller deoxiribos. I deoxiribos ersätts OH-gruppen vid den andra kolatomen med en H-atom, det vill säga den har en syreatom mindre än i ribos.
I båda typer av nukleinsyror innehåller baser av fyra olika typer: två av dem tillhör klassen puriner och två till klassen pyrimidiner. Kvävet som ingår i ringen ger huvudkaraktären till dessa föreningar. Puriner inkluderar adenin (A) och guanin (G), och pyrimidiner inkluderar cytosin (C) och tymin (T) eller uracil (U) (i DNA respektive RNA). Tymin är kemiskt mycket nära uracil (det är 5-metyluracil, det vill säga uracil, där en metylgrupp finns vid den 5:e kolatomen). Purinmolekylen har två ringar, medan pyrimidinmolekylen har en.
Grunder Det är vanligt att beteckna den första bokstaven i deras namn: A, G, T, U och C.

Nukleinsyrorär syror eftersom deras molekyl innehåller fosforsyra.
Figuren visar hur socker, bas och fosforsyra kombineras för att bildas nukleotidmolekyl. Kombinationen av socker med en bas sker med frisättningen av en vattenmolekyl, det vill säga det är en kondensationsreaktion. För bildandet av en nukleotid krävs ytterligare en kondensationsreaktion - mellan socker och fosforsyra.
Diverse nukleotider skiljer sig från varandra i sockerarternas karaktär och de baser som ingår i dem.
Nukleotidernas roll i kroppenär inte begränsad till att fungera som byggstenar för nukleinsyror; några viktiga koenzymer är också nukleotider. Dessa är till exempel adenosintrifosfat (ATP), cykliskt adenosinmonofosfat (cAMP), koenzym A, nikotinamidadenindinukleotid (NAD), ni(NADP) och flavinadenindinukleotid (FAD).
Nukleinsyror är naturliga makromolekylära föreningar (polynukleotider) som spelar en stor roll vid lagring och överföring av ärftlig information i levande organismer.
Molekylvikten hos nukleinsyror kan variera från hundratusentals till tiotals miljarder. De upptäcktes och isolerades från cellkärnor redan på 1800-talet, men deras biologiska roll klargjordes först under andra hälften av 1900-talet.
Nukleotidens sammansättning - den strukturella enheten av nukleinsyror - innehåller tre komponenter:
1) kvävehaltig bas - pyrimidin eller purin
Pyrimidinbaser- pyrimidinderivat som ingår i nukleinsyror:uracil, tymin, cytosin.
För baser som innehåller –OH-gruppen är en mobil jämvikt av strukturella isomerer karakteristisk, på grund av överföringen av en proton från syre till kväve och vice versa:
Purinbaser- purinderivat som ingår i nukleinsyror: adenin, guanin.
Guanin finns som två strukturella isomerer:
2) monosackarid
Ribos och 2-deoxiribos avser monosackarider som innehåller fem kolatomer. De ingår i sammansättningen av nukleinsyror i cykliska β-former:
3) fosforsyrarest
DNA och RNA
Beroende på vilken monosackarid som finns i polynukleotidens strukturella enhet - ribose eller 2-deoxiribos, skilja på
· ribonukleinsyror(RNA) och
· deoxiribonukleinsyror(DNA)
Huvudsträngen (socker-fosfat) av RNA innehåller rester ribose och i DNA 2-deoxiribos.
Nukleotidenheter av DNA-makromolekyler kan innehålla adenin, guanin, cytosin och tymin. Sammansättningen av RNA skiljer sig i att istället för tymin närvarande uracil.
Molekylvikten hos DNA når tiotals miljoner amu. Dessa är de längsta kända makromolekylerna. Molekylvikten för RNA är mycket lägre (från flera hundra till tiotusentals). DNA finns främst i cellkärnorna, RNA - i cellers ribosomer och protoplasma.
När strukturen av nukleinsyror beskrivs, beaktas olika nivåer av organisering av makromolekyler:primär och sekundär strukturera.
· Primär struktur nukleinsyror är nukleotidsammansättningen och en viss sekvens av nukleotidenheter i polymerkedjan.
Till exempel:
I den förkortade enbokstavsbeteckningen skrivs denna struktur som
...– A – G – C –...
· Under sekundär struktur nukleinsyror förstår de rumsligt ordnade formerna av polynukleotidkedjor.
Sekundär struktur av DNAbestår av två parallella ogrenade polynukleotidkedjor vridna runt en gemensam axel till en dubbelhelix.
Denna tredimensionella struktur hålls av många vätebindningar som bildas av kvävehaltiga baser riktade inåt spiralen.Vätebindningar uppstår mellan purinbasen i en kedja och pyrimidinbasen i den andra kedjan. Dessa baser utgör komplementära par (från lat. komplementum- tillägg).
Bildandet av vätebindningar mellan komplementära baspar beror på deras rumsliga överensstämmelse.
Pyrimidinbasen är komplementär till purinbasen:
Vätebindningar mellan andra baspar tillåter dem inte att passa in i dubbelspiralstrukturen. Således,
TYMIN (T) är komplementär till Adenin (A),
CYTOSIN (C) är komplementär till GUANIN (G).
Baskomplementaritet avgörkedjekomplementariteti DNA-molekyler.
Komplementariteten hos polynukleotidkedjor fungerar som den kemiska grunden för DNA:s huvudfunktion - lagring och överföring av ärftliga egenskaper.
DNA:s förmåga att inte bara lagra utan också använda genetisk information bestäms av dess följande egenskaper:
DNA-molekyler är kapabla till replikation (fördubbling), d.v.s. kan möjliggöra syntesen av andra DNA-molekyler som är identiska med de ursprungliga, eftersom sekvensen av baser i en av kedjorna i dubbelhelixen styr deras placering i den andra kedjan.
DNA-molekyler kan styra syntesen av proteiner specifika för organismer av en given art på ett helt exakt och bestämt sätt.
Sekundär struktur av RNA
Till skillnad från DNA består RNA-molekyler av en enda polynukleotidkedja och har inte en strikt definierad rumslig form (RNA:s sekundära struktur beror på deras biologiska funktioner).
Huvudrollen för RNA är direkt deltagande i proteinbiosyntes.
Tre typer av cellulärt RNA är kända, som skiljer sig åt i deras placering i cellen, sammansättning, storlek och egenskaper som bestämmer deras specifika roll i bildandet av proteinmakromolekyler:
informations-(matris)RNA:n överför information kodad i DNA om proteinets struktur från cellkärnan till ribosomerna, där proteinsyntesen utförs;
transport-RNA samlar aminosyror i cellens cytoplasma och överför dem till ribosomen; RNA-molekyler av denna typ "lär sig" från motsvarande sektioner av budbärar-RNA-kedjan vilka aminosyror som bör delta i proteinsyntesen;
Ribosomala RNA tillhandahåller proteinsyntes av en viss struktur och läser information från informations-RNA (matris).
är komplexa monomerer från vilka heteropolymermolekyler är sammansatta. DNA och RNA. Fria nukleotider är involverade i livets signal- och energiprocesser. DNA-nukleotider och RNA-nukleotider har en gemensam strukturplan, men skiljer sig i strukturen av pentossockret. DNA-nukleotider använder sockret deoxiribos, medan RNA-nukleotider använder ribos.
Strukturen av en nukleotid
Varje nukleotid kan delas in i 3 delar:
1. En kolhydrat är ett femledat pentossocker (ribos eller deoxiribos).
2. Fosforrest (fosfat) är resten av fosforsyra.
3. En kvävehaltig bas är en förening i vilken det finns många kväveatomer. I nukleinsyror används endast 5 typer av kvävehaltiga baser: Adenin, Tymin, Guanin, Cytosin, Uracil. Det finns 4 typer i DNA: adenin, tymin, guanin, cytosin. I RNA finns även 4 typer: Adenin, Uracil, Guanin, Cytosin.Det är lätt att se att i RNA är tymin ersatt av Uracil jämfört med DNA.
Den allmänna strukturformeln för pentos (ribos eller deoxiribos), vars molekyler bildar "skelettet" av nukleinsyror:
Om X ersätts med H (X = H), erhålls deoxiribonukleosider; om X ersätts med OH (X = OH), erhålls ribonukleosider. Om vi ersätter en kvävebas (purin eller pyrimidin) istället för R, så får vi en specifik nukleotid.
Det är viktigt att vara uppmärksam på de positioner av kolatomer i pentos, som är betecknade som 3" och 5". Numreringen av kolatomer börjar från syreatomen överst och går medurs. Den sista kolatomen (5") erhålls, som ligger utanför pentosringen och bildar, man kan säga, en "svans" av pentosen. Så när man bygger en kedja av nukleotider kan enzymet bara fästa en ny nukleotid till kol 3" och till ingen annan . Därför kan 5"-änden av nukleotidkedjan aldrig fortsätta, bara 3"-änden kan förlängas.



Jämför en nukleotid för RNA med en nukleotid för DNA.
Försök att ta reda på vilken nukleotid det är i denna representation:

ATP - fri nukleotid
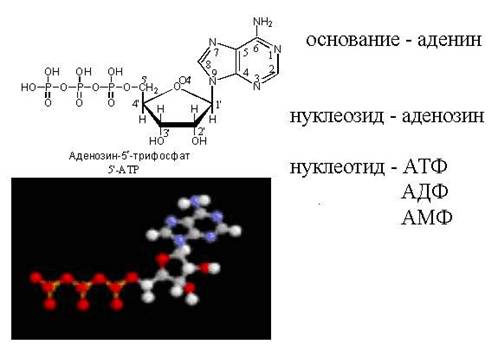
cAMP - "loopback" ATP-molekyl

Diagram över nukleotidstrukturen

Observera att en aktiverad nukleotid som kan bygga upp en DNA- eller RNA-kedja har en "trifosfatsvans". Det är med denna "energimättade" svans som den kan ansluta sig till den redan existerande kedjan av den växande nukleinsyran. Fosfatsvansen sitter på kol 5, så att kolpositionen redan är upptagen av fosfater och är tänkt att fästas. Vad ska man fästa den på? Endast till kolet i position 3". När den väl är fäst kommer denna nukleotid själv att bli ett mål för fästningen av nästa nukleotid. Den "mottagande sidan" tillhandahåller kolet i position 3", och den "inkommande sidan" klamrar sig fast vid den med en fosfatsvans belägen vid position 5". I allmänhet växer kedjan från 3"-sidan.
Förlängning av DNA-nukleotidkedjan
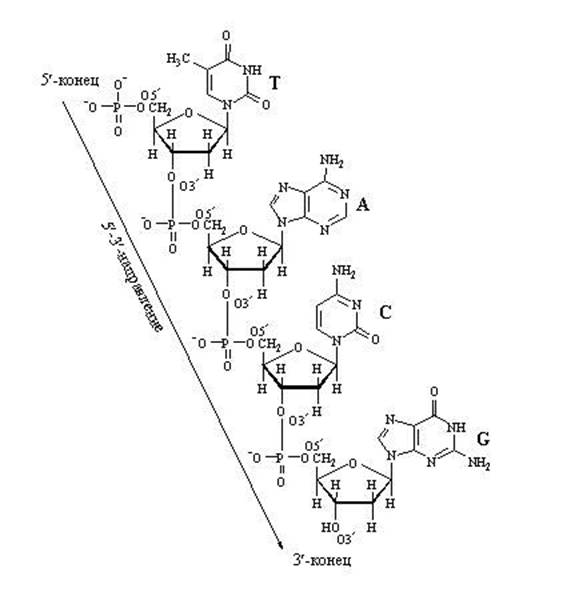
Kedjetillväxt på grund av "längsgående" bindningar mellan nukleotider kan bara gå i en riktning: från 5" ⇒ till 3", eftersom En ny nukleotid kan bara läggas till 3'-änden av kedjan, inte till 5'-änden.
Par av nukleotider sammankopplade genom "korsande" komplementära bindningar av deras kvävehaltiga baser
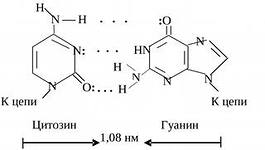
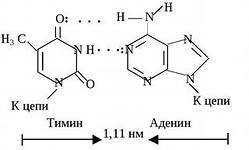
Sektion av DNA-dubbelhelixen

Hitta tecken på antiparallellism av två DNA-strängar.
Hitta nukleotidpar med dubbel- och trippelkomplementära bindningar.