Vilka organ bildas av groddskikten. groddskiktsteori. Derivat av groddskikten. Historien om utvecklingen av teorin om groddlager
Groddlager är en grundläggande term inom embryologi. De betecknar fosterkroppens skikt i ett tidigt skede, i de flesta fall är dessa skikt av epitel.
Groddskikten klassificeras vanligtvis i tre typer:
Ektoderm - det yttre arket, som också kallas epiblast eller hudkänsligt lager;
Endoderm är det inre lagret av celler. Det kan också kallas ett hypoblast eller entero-körtelark;
Mellanskikt (mesoderm eller mesoblast).
Germinala ark (beroende på deras placering kännetecknas de av vissa cellegenskaper. Således består det yttre lagret av embryot av ljusa och höga celler, som i struktur liknar cylindriskt epitel. Det inre bladet består i de flesta fall av stora celler som är fyllda med specifika äggulaplattor. De har ett tillplattat utseende, vilket gör att de ser ut
Mesodermen i det första stadiet består av spindelformade och stellatiska celler. De bildar senare epitelskiktet. Jag måste säga att många forskare tror att mesodermen är de mellersta groddskikten, som inte är ett oberoende skikt av celler.
Groddskikten har till en början formen av en ihålig formation, som kallas blastodermal vesikel. Vid en av dess poler samlas en grupp celler, som kallas cellmassa. Det ger upphov till primärtarmen (endoderm).
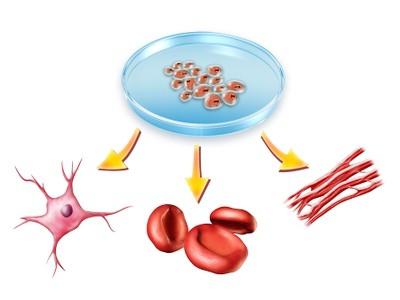
Det bör sägas att olika organ bildas av embryonala ark. Således uppstår nervsystemet från ektodermen, matsmältningsröret kommer från endodermen och skelettet och musklerna kommer från mesodermen.
Det bör också noteras att speciella embryonala membran bildas under embryogenes. De är tillfälliga, deltar inte i bildandet av organ och existerar endast under embryonal utveckling. Varje klass har vissa funktioner i bildningen och strukturen av dessa skal.

Med utvecklingen av embryologi började de bestämma likheten mellan embryon, som först beskrevs av K.M. Baer 1828. Lite senare identifierade Charles Darwin huvudorsaken till likheten mellan embryon från alla organismer - deras gemensamma ursprung. Severov, å andra sidan, hävdade att de vanliga tecknen på embryon är förknippade med evolution, som i de flesta fall fortsätter genom anabolism.
När man jämförde huvudstadierna i utvecklingen av embryon av olika klasser och djurarter, hittades vissa egenskaper som gjorde det möjligt att formulera lagen om embryonal likhet. Huvudbestämmelserna i denna lag var att embryon från organismer av samma typ i de tidiga stadierna av deras utveckling är mycket lika. Därefter kännetecknas embryot av fler och fler individuella egenskaper som indikerar dess tillhörighet till motsvarande släkte och art. Samtidigt separeras embryona från representanter av samma typ alltmer från varandra, och deras primära likhet spåras inte längre.
groddlager(lat. embryonal folia), groddlager, lager av embryots kropp hos flercelliga djur, bildade under gastrulation och ger upphov till olika organ och vävnader. I de flesta organismer bildas tre groddlager: det yttre är ektodermen, det inre är endodermen och det mellersta mesodermen.
Derivat av ektodermen utför huvudsakligen integumentära och sensoriska funktioner, derivat av endodermen - funktionerna för näring och andning, och derivat av mesodermen - kopplingar mellan delar av embryot, motoriska, stödjande och trofiska funktioner.
Samma groddlager hos representanter för olika klasser av ryggradsdjur har samma egenskaper, d.v.s. groddlager är homologa formationer och deras närvaro bekräftar positionen för enheten i djurvärldens ursprung. Groddlager bildas i embryon av alla större klasser av ryggradsdjur, d.v.s. är universellt fördelade.
Groddskiktet är ett skikt av celler som intar en viss position. Men det kan inte betraktas endast från topografiska positioner. Groddskiktet är en samling celler som har vissa utvecklingstendenser. Ett klart definierat, om än ganska brett, spektrum av utvecklingspotentialer bestäms (bestäms) slutligen vid slutet av gastrulationen. Således utvecklas varje groddlager i en given riktning, deltar i uppkomsten av rudimenten hos vissa organ. I hela djurriket kommer enskilda organ och vävnader från samma groddlager. Från ektodermen bildas neuralröret och integumentära epitelet, från endodermen - tarmepitelet, från mesodermen - muskel och bindväv, epitelet i njurarna, könskörtlarna och serösa håligheter. Från mesodermen och den kraniala delen av ektodermen vräkas celler, som fyller utrymmet mellan arken och bildar mesenkymet. Mesenkymala celler bildar syncytium: de är anslutna till varandra genom cytoplasmatiska processer. Mesenkymet bildar bindväven. Varje enskilt groddlager är inte en autonom formation, det är en del av helheten. Groddskikten kan endast särskiljas genom att interagera med varandra och vara under påverkan av de integrerande influenserna från embryot som helhet. En bra illustration av sådan interaktion och ömsesidig påverkan är experiment på tidig amfibiegastrula, enligt vilka cellmaterialet i ekto-, ento- och mesoderm kan tvingas att radikalt ändra vägen för dess utveckling, för att delta i bildandet av organ som är helt okaraktäristiska för denna broschyr. Detta tyder på att i början av gastrulation är ödet för cellmaterialet i varje groddlager strikt sett ännu inte förutbestämt. Utvecklingen och differentieringen av varje blad, deras organogenetiska specificitet, beror på den ömsesidiga påverkan av delarna av hela embryot och är endast möjlig med normal integration.
62. Histo- och organogenes. Processen för neurulation. Axiella organ och deras bildning. mesoderm differentiering. Derivatorgan från ryggradsdjurs embryon.
Histogenes(från andra grekiska ἱστός - vävnad + γένεσις - utbildning, utveckling) - en uppsättning processer som leder till bildandet och återställandet av vävnader under individuell utveckling (ontogenes). Ett eller annat groddlager är involverat i bildandet av en viss typ av vävnad. Till exempel utvecklas muskelvävnad från mesoderm, nervvävnad från ektoderm etc. I vissa fall kan vävnader av samma typ ha ett annat ursprung, till exempel är hudens epitel ektodermalt och det absorberande tarmepitelet är endodermalt i ursprung.
Organogenes- det sista stadiet av embryonal individuell utveckling, som föregås av befruktning, krossning, blastulation och gastrulation.
Vid organogenes, neurulation, histogenes och organogenes.
I neurulationsprocessen bildas en neurula, i vilken mesodermen läggs, bestående av tre groddlager (det tredje lagret av mesodermen delas i segmenterade parade strukturer - somiter) och det axiella komplexet av organ - neuralröret, korda och tarm. Cellerna i det axiella komplexet av organ påverkar varandra ömsesidigt. Denna ömsesidiga påverkan kallas embryonal induktion.
I processen med histogenes bildas kroppsvävnader. Från ektodermen bildas nervvävnad och hudens epidermis med hudkörtlar, från vilka nervsystemet, sensoriska organ och epidermis därefter utvecklas. Från endodermen bildas en notokord och epitelvävnad, från vilken slemhinnor, lungor, kapillärer och körtlar (förutom genital och hud) därefter bildas. Mesodermen producerar muskler och bindväv. ODS, blod, hjärta, njurar och könskörtlar bildas av muskelvävnad.
Neurulation- bildandet av nervplattan och dess stängning i nervröret i processen för embryonal utveckling av chordater.
Neurulation är ett av de viktigaste stadierna av ontogeni. Ett embryo i neurulationsstadiet kallas en neurula.
Utvecklingen av neuralröret i anterior-posterior riktning styrs av speciella ämnen - morfogen (de bestämmer vilka av ändarna som kommer att bli hjärnan), och den genetiska informationen om detta är inbäddad i de så kallade homeotiska eller homeotiska generna.
Till exempel kan morfogen retinsyra, med en ökning av dess koncentration, förvandla rombomerer (segment av nervröret i den bakre delen av hjärnan) av en typ till en annan.
Neurulation i lansetter är tillväxten av åsar från ektodermen över ett lager av celler som blir neurala plattan.
Neurulation i det stratifierade epitelet - cellerna i båda skikten går ner under ektodermen blandat och divergerar centrifugalt och bildar ett neuralrör.
Neurulation i ett enskiktigt epitel:
Schizocoelous typ (i teleosts) - liknar stratifierad epitelneurulation, förutom att cellerna i ett lager går ner.
Hos fåglar och däggdjur invaginerar neuralplattan inåt och sluter sig in i nervröret.
Hos fåglar och däggdjur, under neurulation, utskjutande delar av nervplattan kallas neurala veck, är stängda längs neuralrörets hela längd ojämnt.
Vanligtvis stängs mitten av neuralröret först, och sedan går förslutningen till båda ändarna, vilket resulterar i två öppna sektioner - de främre och bakre neuroporerna.
Hos människor är förslutningen av neuralröret mer komplex. Den dorsala sektionen stänger först, från bröstkorgen till ländryggen, den andra - området från pannan till kronan, den tredje - framsidan, går i en riktning, till neurokraniet, den fjärde - området från baksidan av huvudet till slutet av livmoderhalsen, den sista, femte - den sakrala sektionen, går också i en riktning, bort från svanskotan.
När den andra sektionen inte är stängd, hittas en dödlig medfödd defekt - anencefali. Fostret utvecklar ingen hjärna.
När den femte sektionen inte är stängd hittas en medfödd defekt som kan korrigeras - spina bifida eller Spinabifida. Beroende på svårighetsgraden är ryggmärgsbråck uppdelad i flera undertyper.
Vid neurulation bildas neuralröret.
I tvärsnitt, omedelbart efter bildandet, kan tre lager urskiljas i den, från insidan till utsidan:
Ependymal - pseudo-stratifierat lager som innehåller rudimentära celler.
Mantelzonen innehåller migrerande, prolifererande celler som kommer ut från det ependymala lagret.
Den yttre marginalzonen är det lager där nervfibrer bildas.
Det finns 4 axiell kropp: notokord, neuralrör, tarmrör och mesoderm.
Oavsett djurart, omvandlas de celler som migrerar genom regionen av den dorsala läppen av blastoporen därefter till en notokord, och genom regionen av de laterala (laterala) läpparna av blastopore in i det tredje groddskiktet - mesodermen. I högre kordater (fåglar och däggdjur), på grund av invandringen av germinala sköldceller, bildas inte blastoporen under gastrulation. Celler som migrerade genom ryggläppen på blastoporen bildar en korda, en tät cellsträng som ligger längs embryots mittlinje mellan ektodermen och endodermen. Under dess inflytande börjar nervröret bildas i det yttre groddskiktet, och först till sist bildar endodermen tarmröret.
Differentiering (lat. differens. skillnad) av mesodermen börjar i slutet av 3:e utvecklingsveckan. Mesenkymet uppstår från mesodermen.
Den dorsala delen av mesodermen, som ligger på sidorna av kordan, är uppdelad i kroppssegment - somiter, från vilka ben och brosk, tvärstrimmiga skelettmuskler och hud utvecklas (bild 134).
Från den ventrala icke-segmenterade delen av mesodermen - med planchnotomen bildas två plattor: splanchnopleura och somatopleura, från vilka mesotelet i de serösa membranen utvecklas, och utrymmet mellan dem förvandlas till kroppshåligheter, matsmältningsröret, blodkroppar, glatt muskelvävnad, blod- och lymfkärl, bindväv, tvärstrimmig hjärtmuskelvävnad, binjurebarken och epitelkönskörtlar.
Derivat av groddskikten. Ektodermen ger upphov till det yttre integumentet, det centrala nervsystemet och den sista delen av matsmältningskanalen. Från endodermen bildas notokorden, mittsektionen av matsmältningsröret och andningsorganen. Från mesodermen bildas muskuloskeletala, kardiovaskulära och genitourinära systemen.
Upprätta en överensstämmelse mellan människokroppens struktur och det groddlager som den bildades av.
Skriv ner siffrorna som svar, ordna dem i den ordning som motsvarar bokstäverna:
| A | B | PÅ | G | D |
Förklaring.
De viktigaste ektodermala derivaten är neuralröret, nervkammen och alla nervceller som bildas av dem. De sinnesorgan som överför information om syn, ljud, lukt och andra stimuli till nervsystemet utvecklas också från ektodermala anlage. Till exempel bildas ögats näthinna som en utväxt av hjärnan och är därför ett derivat av neuralröret, medan luktceller differentierar sig direkt från det ektodermala epitelet i näshålan. Smärtreceptorer är av ektodermalt ursprung.
Ektoderm: smärtreceptorer, hårfäste, nagelplattor. Mesoderm: lymf och blod, fettvävnad.
Svar: 11221.
Svar: 11221
Källa: Unified State Examination in Biology 2013-05-30. huvudvåg. Sibirien. Alternativ 2.
Sadi 11.06.2017 13:49
I svaret på denna uppgift står det skrivet att lungorna bildas av mesodermen, och i Uppgift 8 nr 13837 sägs det från endodermen.
Natalya Evgenievna Bashtannik
Observera att lungans epitel är endodermen.
Grunden för ett visst organ bildas initialt från ett specifikt groddlager, men sedan blir organet mer komplext, och som ett resultat deltar två eller tre groddlager i dess bildande.
Lungan är inte bara ett epitel, det är också bronkioler och anslutande filmer ... allt detta bildas från mesenkymet, och tyvärr övervägs inte denna kunskap av kompilatorerna i Unified State Examination :(
Utrymmet mellan de utvecklande bronkerna är fyllt med mellanliggande mesenkym. Mesenkymet, som är en lös vävnad som tätt täcker de utvecklande endodermala tubulära formationerna, börjar differentiera vid roten av lungorna under den tredje månaden. Härifrån fortsätter differentieringen i perifer riktning med separata grenar av bronkerna. Först uppträder de båda huvudbronkernas broskringar, och broskplattorna i de återstående bronkerna särskiljer sig gradvis. Ungefär samtidigt bildas muskelceller och bindvävens första kollagenfibrer. Från det mesodermala materialet uppstår interlobulärt och intersegmentalt septalmesenkym och subserös bindväv i lungfilmen. Elastiska fibrer börjar dyka upp i den fjärde månaden. Deras huvudsakliga utveckling sker dock, liksom utvecklingen av broskplattor i bronkernas väggar, endast under andra hälften av den intrauterina utvecklingen.
"MORDOVA STATE UNIVERSITY uppkallad efter A.I. N. P. OGAREVA»
Institutionen för biologi
Institutionen för genetik
på ämnet: groddlager
Genomförd av: 3:e årsstudent
specialitet "biologi"
Introduktion
1. Strukturen av groddskikten
2. Historia om utvecklingen av teorin om groddlager
3. Bildning av groddlager
4. Ursprung och evolutionär betydelse för groddskikten
5. Bestämmelser i teorin om groddlager och invändningar mot denna teori
Slutsats
Litteratur
Introduktion
Tillsammans med möjligheten att tolka groddskikten utifrån deras fylogenetiska betydelse, är det viktigt att fastställa vilken roll de spelar för individuell utveckling. Groddskikten är de första organiserade grupperna av celler i embryot, som tydligt skiljer sig från varandra genom sina egenskaper och relationer. Det faktum att dessa förhållanden i princip är desamma i alla ryggradsdjurs embryon tyder starkt på ett gemensamt ursprung och liknande ärftlighet hos de olika medlemmarna av denna stora grupp djur.
Man kan tro att i dessa groddlager, för första gången, börjar skillnader av olika klasser att skapas ovanför den allmänna planen för kroppsstrukturen, karakteristisk för alla ryggradsdjur.
Bildandet av groddlager avslutar perioden då den huvudsakliga utvecklingsprocessen bara är en ökning av antalet celler, och perioden för differentiering och specialisering av celler börjar. Differentiering sker i groddskikten innan vi kan se tecken på det med någon av våra mikroskopiska metoder. I ett blad som har ett helt enhetligt utseende uppstår ständigt lokaliserade grupper av celler med olika potential för vidareutveckling.
Olika strukturer uppstår från groddskiktet. Samtidigt är inga synliga förändringar omärkliga i groddskiktet, på grund av vilka de uppstår. Nyligen genomförda experimentella studier indikerar hur tidigt denna osynliga differentiering föregår den synliga morfologiska lokaliseringen av cellgrupper, som vi lätt känner igen som rudimentet för det definitiva organet.
1. Strukturen av groddskikten
Groddskikten består av cellmaterial som används för utveckling av olika organ och vävnader. I sin struktur skiljer sig cellerna i olika groddlager från varandra; endodermceller är alltid större och mindre regelbundna än ektodermala celler. Endodermen kännetecknas av egenskaperna hos det framtida bokmärket, som har trofisk betydelse. Ektodermen stannar kvar på ytan och har initialt ett skyddsvärde. Till skillnad från endodermen består den av regelbundet arrangerade celler med en mer enhetlig form. Gastrulation leder till en märkbar skillnad mellan de yttre och inre lagren och groddmaterialet blir heterogent. Processen som leder till uppkomsten av skillnader i ett initialt homogent material kallas differentiering.
Primära organisatörer eller induktorer spelar en viktig roll i differentieringen av cellulärt material. Induktorer är kemikalier som frigörs av grupper av celler och påverkar andra grupper av celler, vilket ändrar deras utvecklingsväg. Som ett resultat av differentiering av groddskikten bildas olika organ och vävnader. Vid studiet av dessa processer hos olika djur fann man att ödet för varje groddlager i alla flercelliga organismer som regel är detsamma.
Således utvecklas hudens epitel, hudkörtlar, många hornderivat, nervsystemet och känselorganen från ektodermen. Från endodermen hos alla djur bildas epitelet i den mellersta delen av tarmkanalen, levern och matsmältningskörtlarna. I chordates bildas också epitelet i andningsvägarna. Blod och lymfa, muskler, bindväv, brosk- och benvävnader, njurepitel, väggen i den sekundära kroppshålan, en del av reproduktionssystemets vävnader utvecklas från mesodermen.
2. Historia om utvecklingen av teorin om groddlager
Groddskiktsteorin är en av de största generaliseringarna av jämförande embryologi under 1800-talet. Groddlagren beskrevs först av X. Pander (1817), som upptäckte att kycklingembryot i vissa utvecklingsstadier består av tre tunna filmer eller lager, vars cellulära karaktär ännu inte var känd. Pander kallade det yttre bladet seröst, det djupaste - slemhinna och det mellanliggande - blod. Dessa observationer bekräftades av K. Baer (1828, 1837), som hittade groddlager hos några andra djur (fiskar, grodor, sköldpaddor). Baer särskiljde två primära lager - animaliskt och vegetativt, som sedan återigen delas in i sekundära groddlager: djurlagret ger hud och muskulärt, och det vegetativa - vaskulärt och slemhinnat. Enligt modern terminologi motsvarar hudskiktet ektodermen, slemskiktet motsvarar endodermen och det muskulära och kärlskiktet motsvarar det parietala och viscerala skiktet av mesodermen. Baers misstag var bara att han beskrev ursprunget till dessa två mesodermala lager hos ryggradsdjur från olika källor. Termerna "ektoderm" och "endoderm" lånades av embryologer från zoologin (så kallades epitelskikten som utgör kroppen av vuxna Cnidarians ännu tidigare). Den cellulära strukturen för kycklingembryots könslager fastställdes av Remak 1855.
Ursprungligen trodde man att groddlager bildas endast under utvecklingen av ryggradsdjur. Men efter arbetet av A. O. Kovalevsky och I. I. Mechnikov, som studerade utvecklingen av nästan alla klasser av ryggradslösa djur, blev det klart att groddlager finns i en eller annan form i alla flercelliga djur. A. O. Kovalevsky (1871) skrev i artikeln "Embryological Studies of Worms and Arthropods" i den sista delen: "Om vi nu jämför utvecklingen av de maskar vi har beskrivit med utvecklingen av andra djur, så är liknelsen mellan groddskikten med de hos ryggradsdjur är särskilt slående för oss, ner till enskilda detaljer; samma två primära blad som spelar en viktig roll i utvecklingen av maskar finns också hos ryggradsdjur; som i vissa, så i andra, uppträder mittbladet först senare. Bladens öden och organens läggning sammanfaller extremt, ända ner till individuella processer.
I. I. Mechnikov upptäckte groddlager hos vissa djur med kraftigt förändrad utveckling och tog för första gången upp frågan om utvecklingen av gastrulationsprocesser.
3. Bildning av groddlager
Groddlager bildas hos djur och människor i en process som kallas gastrulation.
Bland djur urskiljs tvålagers och trelagers taxa. Från och med plattmaskar har djur 3 groddlager: ektoderm (yttre), endoderm (inre) och mesoderm (mitten). Mesoderm finns endast hos trelagersdjur, medan ektoderm och endoderm finns hos tvålagers (svampar, mossor, coelenterater) och trelagersdjur.
Nervsystemet, huden, hudkörtlarna, hudderivat, såsom fjädrar, hår, naglar, klor, fjäll, liksom epitelet i de främre och bakre delarna av matsmältningsröret, och benen i det viscerala skelettet utvecklas från ektoderm i ontogeni.
Tarmslemhinnan bildas av endodermen; endodermen ger näring till embryot; från detta groddlager utvecklas andningsorganen, matsmältningssystemets slemhinnor och matsmältningskörtlarna (lever etc.).
Från mesodermen bildas organen i cirkulations-, utsöndrings- och reproduktionssystemen, de serösa membranen i coelom och inre organ, såväl som benen i det stödjande skelettet och musklerna.
Moderna metoder för att studera den embryonala processen har gjort det möjligt att fastställa att groddskikten inte har betydelsen av ett primitivt organ och inte upprepar något stadium av fylogenetisk utveckling. De bör betraktas som materialet i ett visst komplex av framtida organ som är på samma utvecklingsnivå och är morfologiskt lika. Processen för bildning av groddlager betyder ett visst stadium i utvecklingen av organ, som de allra flesta djur går igenom.
Vanligtvis består varje organ av vävnader som härrör från olika groddlager, men vi klassificerar ett organ som ett derivat av ett eller annat blad, beroende på vad dess huvudsakliga primordium utvecklas från. Således består mellantarmsväggen hos ryggradsdjur av endodermalt epitel och mesodermala glatta muskler i ursprung och ett lager av bindväv. Men eftersom den första rudimenten av mellantarmen bildas från endodermen, och de mesodermala elementen fästs vid den senare, och matsmältningsfunktionen utförs av endodermala epitelet, anses mellantarmen vara ett endodermalt organ.
Närvaron av groddlager, på samma sätt involverade i konstruktionen av kroppen av alla Metazoa, gjorde det möjligt att jämföra utvecklingen av systematiskt avlägsna grupper av djur. För närvarande är det helt enkelt omöjligt att beskriva utvecklingen av något djur utan att nämna groddskikten.
4. Ursprung och evolutionär betydelse för groddskikten
Frågan uppstår om vad som är ursprunget och den evolutionära betydelsen av groddskikten. Enligt E. Haeckel (1874) upprepar de primära groddskikten (ekto- och endoderm) i utvecklingen (rekapitulerar) de primära organen (hud och tarmar) hos den hypotetiska gemensamma förfadern till Metazoa - Gastrea. Av detta följer att groddskikten hos alla djur är homologa. I. I. Mechnikov (1886) fäste också rekapitulationsbetydelse åt groddlagren, men han representerade den gemensamma förfadern till Metazoa i form av Phagocytella. Enligt Mechnikov representeras kinoblasten under utvecklingen av ektodermen, och alla organ som uppstod i evolutionsprocessen från kinoblasten har ett ektodermalt ursprung under individuell utveckling. Utvecklingen av fagocytoblasten skedde i två riktningar. Hos coelenterates epiteliserades det fullständigt och förvandlades till en slemhinna i maghålan; i individuell utveckling representeras den av endodermen. Hos treskiktade djur förvandlades endast den centrala delen av fagocytoblasten till tarmen och representeras i ontogenesen av endodermen, medan den perifera delen gav upphov till vävnader i den inre miljön och representeras i ontogenesen av mesodermen.
5. Bestämmelser i teorin om groddlager och invändningar mot denna teori
Således, i slutet av XIX-talet. den klassiska teorin om groddlager har utvecklats, vars innehåll är följande bestämmelser:
1. Vid ontogenesen av alla flercelliga djur bildas två eller tre groddlager, från vilka alla organ utvecklas.
2. Groddskikten kännetecknas av en viss position i embryots kropp (topografi) och betecknas som ekto-, ento- och mesoderm.
3. Groddskikten är specifika, det vill säga vart och ett av dem ger upphov till strikt definierade primordier, som är lika hos alla djur.
4. Groddskikten rekapitulerar i ontogeni de primära organen hos den gemensamma förfadern till alla Metazoer och är därför homologa.
5. Den ontogenetiska utvecklingen av ett organ från ett eller annat groddlager indikerar dess evolutionära ursprung från det motsvarande primära organet hos förfadern.
Hittills har många fakta samlats som vid första anblicken inte passar in i ramen för den klassiska teorin om groddlager. Därför började det dyka upp uttalanden om att denna teori är föråldrad, befinner sig i kris och behöver revideras. All denna kritik är baserad på en alltför formell anti-evolutionär förståelse av groddlagren.Låt oss överväga några av de viktigaste invändningarna mot teorin om groddlager.
1. Det faktum att mesodermen kan härröra från både ektodermen och endodermen har varit föremål för många meningsskiljaktigheter, och detta ställer tvivel om dess enhet som groddlager. Många författare anser att det är nödvändigt att skilja mellan mesoblasten (entomesoderm) och mesenkymet (ektomesoderm). Men skillnaderna mellan dessa delar av mesodermen är inte så betydande som det verkar vid första anblicken. I former med spiralfragmentering härstammar mesenkymet från mikromerer av den 2:a och 3:e kvartetten, och mesoblasten tillhör den 4:e kvartetten: alla dessa celler är belägna längs kanterna av blastoporen, dvs i gränszonen mellan ekto- och ekto- och endoderm. Migration av mesenkymala element till blastocoel är en del av gastrulation. Det kan också antas att den evolutionära bildningen av fagocytoblasten, vars perifera del representeras av mesodermen, var en lång process, och dess påfyllning på grund av kinoblasten fortsatte under mycket lång tid, vilket återspeglas i ontogeni.
2. Hos vissa djur presenteras groddskikten i en mycket komplicerad form. Hos exempelvis Insekter och Fåglar observeras den så kallade bifasiska eller till och med flerfasiga gastrulationen, som så att säga bryts upp i ett antal självständiga handlingar. Ofta, redan innan bildningen av groddlager är avslutad, börjar organogenesen, organens rudiment är isolerade. Groddskikten är inte tydligt uttryckta. Men denna situation kan lätt förklaras som ett resultat av en sekundär förändring i utvecklingens gång. Vi får inte glömma att alla ontogenetiska processer är föremål för evolution i samma utsträckning som organen hos vuxna djur. Även inom phylum Cnidaria har gastrulation genomgått en avsevärd utveckling, så det är inte förvånande att hos högre djur långt borta från ursprunget till Metazoa har gastrulationsprocesser genomgått sådana djupgående sekundära förändringar. Snarare bör man bli förvånad över att vi fortfarande urskiljer groddlager i dem, om än i modifierad form.
3. När det gäller strikt bestämd klyvning (hos nematoder, annelider, blötdjur, ascidier) representerar redan individuella blastomerer eller grupper av blastomerer rudimenten av vissa organ. Så, i den ringade masken Arenicola, vid scenen av 64 blastomerer, urskiljs den så kallade rosetten, bestående av 4 celler, som är rudimentet för en känslig sultan, på djurpolen, och i ekvatorialzonen finns det 4 grupper av celler, 4 i varje - trochoblaster, från vilka det utvecklar prototroch. Vid den vegetativa polen finns det 7 stora celler rika på äggula - tarmens rudiment, till vilken en cell ligger intill från den framtida ryggsidan, vilket ger upphov till mesodermala teloblaster. Man får intrycket att de senare bildade groddskikten inte har någon självständig betydelse, utan endast är en tillfällig sammanslutning av redan existerande heterogena rudiment.
Denna association av primordia i groddskikten är dock inte tillfällig, utan är historiskt betingad. Så sammansättningen av ektodermen inkluderar rudimenten av endast de organ som utvecklas från den och med icke-deterministisk fragmentering (hud, sensoriska organ, etc.). Dessutom är den tidiga bestämningen av blastomerer också en konsekvens av sekundära förändringar under utvecklingsförloppet - detta är en anpassning som gör att embryot snabbt kan förvandlas till en larv, bestående av några fler celler, men som redan är kapabel att självständigt utföra alla vitala funktioner (förutom, naturligtvis, sexuella).
4. Kritiker av groddskiktsteorin brukar peka på förekomsten av olika undantag, som inkluderar perversion av groddskikten i svampar, frånvaron av tydligt uttryckta skikt hos många plattmaskar, frånvaron av endoderm i de flesta bryozoaner, etc. Vi kommer att överväga alla dessa specifika exempel tillsammans med en detaljerad beskrivning av utvecklingen av dessa djur. Vi noterar bara att förekomsten av alla särskilda avvikelser från den allmänna regeln helt kan förstås ur en evolutionär synvinkel, och orsakerna som orsakade dem är tydliga i de flesta fall. Dessutom observeras dessa avvikelser vanligtvis hos ganska lågt organiserade djur, medan hos högre djur (leddjur, ryggradsdjur), specificiteten hos groddskikten observeras strikt. Detta tyder på att groddskikten i de nedre Metazoerna är mycket labila, medan deras specificitet dök upp senare och fortskrider under evolutionens gång.
5. Vid asexuell reproduktion, olika återställande processer och experimentellt ingripande under utvecklingen, observeras ofta ett brott mot principen om specificitet för groddlager. Så under spirande av Bryozoans och vissa Ascidia ingår inte vävnader av endodermal natur i njurens sammansättning, och tarmen utvecklas från ektodermen. Hos Nemertine Lineus lacteus kan en liten pre-oral del av kroppen skäras av, som inte heller innehåller endodermala organ, och ett helt djur utvecklas från detta fragment.
För att förstå arten av dessa fenomen är det nödvändigt att komma ihåg vad bakterielagrens specificitet är baserad på. I embryogenesen, från varje blad, utvecklas de organ som historiskt sett separerade från sammansättningen av motsvarande cellskikt, dvs. bladens specificitet är baserad på fenomenet rekapitulation. Själva rekapituleringen (som visas av I. I. Shmalgauzen) beror till stor del på det faktum att vissa historiskt etablerade morfogenetiska korrelationer existerar mellan embryots delar. Men under loppet av återhämtningsprocesser och asexuell reproduktion fortsätter utvecklingen inte på basis av gastrula, utan på basis av vävnader hos ett vuxet djur, mellan vilka det finns andra fysiologiska relationer. Groddskikten är uteslutande embryonala formationer och är som sådana frånvarande hos vuxna djur. Därför förlorar bakterielagrens specificitet sin betydelse.
Till detta kan vi lägga till att förmågan till asexuell reproduktion och bredare morfogenetiska förmågor hos vävnader endast är karakteristiska för djur som inte har nått en mycket hög evolutionär nivå, vilket indikerar den progressiva specificiteten hos groddskikten och vävnaderna hos ett vuxet djur.
Den moderna synen på groddskikten uttrycks väl av följande citat från V. N. Beklemishevs "Comparative Anatomy of Invertebrates": "... kinoblasten och fagocytoblasten är huvudskikten i kroppen och djurets direkta organ endast i larverna av coelenterates och svampar och i de mest enkelt arrangerade hydroider, som Protohydra. I alla andra Enterozoa, på grund av koncentrationen av funktioner och integrering av organ, bryts de primära lagren upp i ett antal derivat, som är sammanflätade på ett komplext sätt. På grund av detta, i de överlägsna Metazoa, reduceras de primära lagren till nivån av groddlager; de är inte längre, som sådana, i den vuxna, utan de bevaras i form av primära skikt av embryot, vilket ger upphov till vissa cellulära system, vävnader och elementära organ i den vuxna organismen. Dessa groddlager förblir dock homologa med varandra i alla Metazoa överallt utom vuxna svampar, och behåller samma grundläggande uppsättningar av karakteristiska egenskaper av ömsesidig position och framtida betydelse.
Slutsats
Så, groddskikten är inte ett imaginärt koncept, de existerar verkligen, de manifesterar en viss typ av primär differentiering av cellulärt material under utvecklingen av Metazoa från ägget. Den konstanthet med vilken groddlagren reproduceras i utvecklingen av de allra flesta djur kan endast förklaras av existensen av "historiska traditioner", det vill säga rekapitulation. Men groddskikten ska inte betraktas som något stabilt och oföränderligt; man bör inte glömma eventuella evolutionära transformationer av alla ontogenetiska processer, inklusive utvecklingen av groddlager.
Litteratur
1. Ivanova-Kazas O. M., Krichinskaya E. B. En kurs i jämförande embryologi av ryggradslösa djur. L. Förlag Leningrad. Universitet, 1988.
2. http:///biologia/26-zarodyshevye-listki. html
3. Great Soviet Encyclopedia, TSB
Vad är groddlager eller lager? Vad är meningen med denna term? Artikeln kommer att ge kort information om dessa isolerade grupper av celler som finns i alla embryon från representanter för faunan i ett visst stadium av embryonal utveckling.
Från historien
Redan på 60-talet av 1700-talet observerade den tyske och ryske fysiologen Caspar Friedrich Wolf och beskrev senare bildandet och omvandlingen till tarmröret av ett av groddskikten. För första gången upptäcktes och beskrevs alla tre bakterielagren av Christian Heinrich Pander, akademiker vid Imperial Academy of Sciences i St. Petersburg (1821), naturforskare, embryolog och paleontolog. Han studerade deras struktur och undersökte också kycklingembryot. Dessutom upptäckte akademikern vid samma akademi, Karl Baer, närvaron av groddlager i embryon från andra djur - fiskar, reptiler, amfibier. Tack vare dessa forskares arbete gavs en impuls till studiet av dessa strukturer.
Bildning av groddlager
Zygoten (ett djurs befruktade ägg) börjar dela sig. I ett tidigt skede av embryonal utveckling delar sig celler intensivt genom mitos och bildar en sfärisk struktur - morula och sedan - blastula. Dess skillnad från morulan ligger i det faktum att cellerna (de kallas blastomerer) i detta skede divergerar från centrum till periferin, och den så kallade blastoderm-vesikeln bildas i mitten. Blastula är därför ett enskiktigt embryo.

Efter slutet av denna period av embryonal utveckling av representanter för djurvärlden, kallad krossning, börjar gastrulationsstadiets tur. Skillnaden mellan dessa stadier av ontogenes är kardinal. I det första fallet är det befruktade ägget uppdelat i många blastomerer (mindre celler), utan att ändra i massa och volym. Den huvudsakliga betydelsen av krossning är embryots övergång från en cell till multicellularitet. Gastrulation, som uppstår efter krossning, innebär celldifferentiering. I detta skede uppstår de så kallade groddskikten. Dessa är vissa grupper av celler från vilka vissa vävnader och organ sedan bildas.
Skillnader i groddlager
Embryots struktur vid gastrulationsstadiet och före det visas på bilden nedan. I stadiet efter gastrulation, kallad neurula, bildas neurala plattan, korda rudiment, epitel och tarmar. De bakre och främre delarna av kroppen blir urskiljbara.

Under gastrulation sker, som nämnts ovan, inte bara cellförökning, utan också deras tillväxt och riktade rörelse, vilket sedan leder till en uttalad differentiering. Grupper av relaterade celler kombineras till separata lager av celler, externa och interna. De kallas ektoderm och endoderm.
Svampar och coelenterater (maneter, koraller, ctenoforer) utvecklar endast dessa två groddlager. Hos högre djur bildas tre av dem: den nämnda ektodermen och endodermen, samt mellanbladet - mesodermen.
Deras skillnader ligger främst i funktioner, samt i början av vilka organ och vävnader de ger upphov till. De kommer att diskuteras mer i detalj nedan.

ektoderm
Det yttre lagret av könsceller ansvarar för motoriska, sensoriska och integumentära funktioner. Därefter utvecklas nervsystemets organ från det. Dessutom utvecklas huden och allt som djuren har på sig från ektodermen: skyddande fjäll, klor, naglar, fjädrar, sköldar etc., samt tandemalj.
Detta groddlager hos ryggradsdjur innehåller tre delar: det yttre, såväl som neuralröret och nervkammen. De två sista komponenterna är också kända som neuroektodermen. Det neurala krönet, på förslag av den kanadensiske embryologen Brian Hall, har kallats det fjärde bakterielagret sedan 2000 i många publikationer.
Endoderm
Det groddlager från vilket inre organ delvis bildas. Detta är matsmältningssystemet, inklusive körtlarna (bukspottkörteln, levern). Andningsorgan utvecklas också från endodermen (i fisk, gälar och en simblåsa).
mesoderm
Mellanskiktet av könsceller, karakteristiskt endast för högre djur. Ansvarig för implementering av trofisk- och stödfunktioner. Det utvecklar ben och muskler, brosk, notokord, utsöndringsorgan, såväl som organ i reproduktions- och cirkulationssystemet.
Till sist
Artikeln beskrev kortfattat djurens groddlager, deras funktioner, listade de organ och system som utvecklas från mesoderm, ektoderm, endoderm.
Ett intressant faktum är att det i alla representanter för djurvärlden i de flesta organ finns vävnader från 2-3 av dessa strukturer.