DNA nukleotiidi koostis. Nukleotiidid. Ühend. Struktuur. Nukleiinhapete koostis
Desoksüribonukleiinhapped (DNA) on makromolekul (üks kolmest põhilisest, ülejäänud kaks on RNA ja valgud), mis tagab talletamise, põlvest põlve edasikandmise ja elusorganismide arengu ja funktsioneerimise geneetilise programmi rakendamise. DNA sisaldab teavet erinevat tüüpi RNA ja valkude struktuuri kohta.
Eukarüootsetes rakkudes (loomad, taimed ja seened) leidub DNA-d raku tuumas kromosoomide osana, aga ka mõnes raku organellis (mitokondrid ja plastiidid). Prokarüootsete organismide (bakterid ja arheed) rakkudes on seestpoolt rakumembraani külge kinnitunud ringikujuline või lineaarne DNA molekul, nn nukleoid. Neil ja madalamatel eukarüootidel (näiteks pärmil) on ka väikesed autonoomsed, enamasti ringikujulised DNA molekulid, mida nimetatakse plasmiidideks. Lisaks võivad ühe- või kaheahelalised DNA molekulid moodustada DNA-d sisaldavate viiruste genoomi.
Keemilisest vaatenurgast on DNA pikk polümeerne molekul, mis koosneb korduvatest plokkidest – nukleotiididest. Iga nukleotiid koosneb lämmastiku alusest, suhkrust (desoksüriboos) ja fosfaatrühmast. Nukleotiidide vahelised sidemed ahelas moodustuvad desoksüriboosist ja fosfaatrühmast (fosfodiestersidemed). Enamikul juhtudest (välja arvatud mõned üheahelalist DNA-d sisaldavad viirused) koosneb DNA makromolekul kahest ahelast, mis on üksteise suhtes orienteeritud lämmastikualuste abil. See kaheahelaline molekul on spiraalne. Üldiselt nimetatakse DNA molekuli struktuuri "topeltspiraaliks".
DNA struktuuri dešifreerimine (1953) oli üks pöördepunkte bioloogia ajaloos. Francis Crick, James Watson ja Maurice Wilkins pälvisid füsioloogia või meditsiini Nobeli preemia 1962. aastal nende silmapaistva panuse eest sellesse avastusse. Rosalind Franklin, kes sai röntgenipildid, ilma milleta poleks Watson ja Crick saanud järeldusi teha. DNA struktuuri, suri 1958. aastal vähki ja Nobeli preemiat ei anta postuumselt.
Ribonukleiinhapped (RNA) on üks kolmest peamisest makromolekulist (ülejäänud kaks on DNA ja valgud), mida leidub kõigi elusorganismide rakkudes.
Nii nagu DNA (desoksüribonukleiinhape), koosneb RNA pikast ahelast, milles iga lüli nimetatakse nukleotiidiks. Iga nukleotiid koosneb lämmastiku alusest, riboossuhkrust ja fosfaatrühmast. Nukleotiidide järjestus võimaldab RNA-l kodeerida geneetilist teavet. Kõik rakulised organismid kasutavad valgusünteesi programmeerimiseks RNA-d (mRNA).
Rakuline RNA moodustub protsessis, mida nimetatakse transkriptsiooniks, st RNA sünteesil DNA matriitsil, mida viivad läbi spetsiaalsed ensüümid - RNA polümeraasid. Messenger RNA-d (mRNA-d) osalevad seejärel protsessis, mida nimetatakse translatsiooniks. Translatsioon on valgu süntees mRNA matriitsil ribosoomide osalusel. Teised RNA-d läbivad pärast transkriptsiooni keemilisi modifikatsioone ning pärast sekundaarsete ja tertsiaarsete struktuuride moodustumist täidavad nad funktsioone, mis sõltuvad RNA tüübist.
Üheahelalisi RNA-sid iseloomustavad mitmesugused ruumilised struktuurid, milles mõned sama ahela nukleotiidid on omavahel paaris. Mõned väga struktureeritud RNA-d osalevad rakuvalgu sünteesis, näiteks ülekande-RNA-d tunnevad ära koodoneid ja toimetavad vastavad aminohapped valgusünteesi kohta, samas kui ribosomaalsed RNA-d on ribosoomide struktuurne ja katalüütiline alus.
Kuid RNA funktsioonid tänapäevastes rakkudes ei piirdu ainult nende rolliga translatsioonis. Seega osalevad väikesed tuuma-RNA-d eukarüootsete messenger-RNA-de splaissimises ja muudes protsessides.
Lisaks sellele, et RNA molekulid on osa mõnedest ensüümidest (näiteks telomeraas), on mõnel RNA-l ka oma ensümaatiline aktiivsus: võime teha teistes RNA molekulides katkestusi või vastupidi kahte RNA fragmenti "liimida". Selliseid RNA-sid nimetatakse ribosüümideks.
Paljude viiruste genoomid koosnevad RNA-st, see tähendab, et see mängib neis rolli, mida DNA mängib kõrgemates organismides. Lähtudes RNA funktsioonide mitmekesisusest rakus, püstitati hüpotees, mille kohaselt on RNA esimene molekul, mis oli võimeline prebioloogilistes süsteemides isepaljunema.
DNA ja RNA vahel on kolm peamist erinevust:
- 1. DNA sisaldab suhkru desoksüriboosi, RNA sisaldab riboosi, millel on desoksüriboosiga võrreldes täiendav hüdroksüülrühm. See rühm suurendab molekuli hüdrolüüsi tõenäosust, see tähendab, et see vähendab RNA molekuli stabiilsust.
- 2. Adeniiniga komplementaarne nukleotiid RNA-s ei ole tümiin, nagu DNA-s, vaid uratsiil on tümiini metüleerimata vorm.
- 3. DNA eksisteerib kaksikheeliksi kujul, mis koosneb kahest eraldiseisvast molekulist. RNA molekulid on keskmiselt palju lühemad ja valdavalt üheahelalised.
Bioloogiliselt aktiivsete RNA molekulide, sh tRNA, rRNA, snRNA ja teiste valke mittekodeerivate molekulide struktuurianalüüs näitas, et need ei koosne ühest pikast heeliksist, vaid arvukatest lühikestest spiraalidest, mis paiknevad üksteise lähedal ja moodustavad midagi sarnast valgu tertsiaarne struktuur. Selle tulemusena võib RNA katalüüsida keemilisi reaktsioone, näiteks ribosoomi peptidüültransferaasi tsenter, mis osaleb valkude peptiidsideme moodustumisel, koosneb täielikult RNA-st.
1944. aastaks avastasid O. Avery ja tema kolleegid K. McLeod ja M. McCarthy DNA transformeeriva aktiivsuse pneumokokkides. Need autorid jätkasid Griffithi tööd, kes kirjeldas transformatsiooni (pärilike tunnuste ülekandumise) fenomeni bakterites. O. Avery, K. McLeod, M. McCarthy näitasid, et valkude, polüsahhariidide ja RNA eemaldamisel ei häirita bakterite transformatsiooni ning kui indutseeriv aine puutub kokku ensüümi desoksüribonukleaasiga, siis transformeeriv aktiivsus kaob.
Nendes katsetes demonstreeriti esimest korda DNA molekuli geneetilist rolli. 1952. aastal kinnitasid A. Hershey ja M. Chase DNA molekuli geneetilist rolli T2 bakteriofaagiga tehtud katsetes. Märgistades selle valgu radioaktiivse väävliga ja selle DNA radioaktiivse fosforiga, nakatasid nad selle bakteriaalse viirusega E. coli. Faagi järglastest leiti suur kogus radioaktiivset fosforit ja ainult jälgi S. Sellest järeldub, et DNA, mitte faagi valk tungis bakterisse ja kandus seejärel pärast replikatsiooni faagi järglastele. .
DNA nukleotiidi struktuur. Nukleotiidide tüübid.
Nukleotiid DNA koosneb
Lämmastikalus (4 tüüpi DNA-s: adeniin, tümiin, tsütosiin, guaniin)
Monosuhkru desoksüriboos
Fosforhappe
nukleotiidi molekul koosneb kolmest osast - viie süsinikusisaldusega suhkrust, lämmastikalusest ja fosforhappest.
Suhkur sisaldub nukleotiidide koostis, sisaldab viit süsinikuaatomit, see tähendab, et see on pentoos. Sõltuvalt nukleotiidis sisalduva pentoosi tüübist on kahte tüüpi nukleiinhappeid - ribonukleiinhapped (RNA), mis sisaldavad riboosi, ja desoksüribonukleiinhapped (DNA), mis sisaldavad desoksüriboosi. Deoksüriboosis asendatakse OH-rühm 2. süsinikuaatomi juures H-aatomiga, see tähendab, et selles on üks hapnikuaatom vähem kui riboosis.
Mõlemas nukleiinhapete tüübid sisaldab nelja erinevat tüüpi aluseid: kaks neist kuuluvad puriinide klassi ja kaks pürimidiinide klassi. Rõngas sisalduv lämmastik annab nendele ühenditele peategelase. Puriinide hulka kuuluvad adeniin (A) ja guaniin (G) ning pürimidiinid tsütosiin (C) ja tümiin (T) või uratsiil (U) (vastavalt DNA-s või RNA-s). Tümiin on keemiliselt väga lähedane uratsiilile (see on 5-metüüluratsiil, st uratsiil, milles 5. süsinikuaatomi juures on metüülrühm). Puriini molekulil on kaks tsüklit, pürimidiini molekulil aga üks.
Nukleotiidid on omavahel seotud tugeva kovalentse sidemega ühe nukleotiidi suhkru ja teise fosforhappe kaudu. Selgub polünukleotiidne ahel. Ühes otsas on vaba fosforhape (5'-ots), teises on vaba suhkur (3'-ots). (DNA polümeraas saab lisada uusi nukleotiide ainult 3'-otsa.)
Kaks polünukleotiidahelat on omavahel ühendatud nõrkade vesiniksidemetega lämmastikualuste vahel. Seal on 2 reeglit:
komplementaarsuse printsiip: tümiin on alati vastand adeniinile, guaniin alati vastupidine tsütosiinile (nad ühtivad üksteisega vesiniksidemete kujul ja arvult - A ja G vahel on kaks sidet ning C ja G vahel 3).
antiparallelismi põhimõte: kus ühel polünukleotiidahelal on 5'-ots, teisel 3'-ots ja vastupidi.
Selgub kahekordne kett DNA.
Ta keerdub sisse kaksikheeliks, ühe spiraali pöörde pikkus on 3,4 nm, sisaldab 10 paari nukleotiide. Lämmastikalused (geneetilise informatsiooni hoidjad) on spiraali sees, kaitstud.
Nukleiinhapped, nagu valgud, on eluks hädavajalikud. Nad esindavad kõigi elusorganismide geneetilist materjali kuni lihtsaimate viirusteni välja. Nimetus "nukleiinhapped" peegeldab asjaolu, et need paiknevad peamiselt tuumas (tuum - tuum). Nukleiinhapete spetsiifilise värvimise korral on tuumad valgusmikroskoobis väga selgelt nähtavad.
DNA struktuuri väljaselgitamine(desoksüribonukleiinhape) - üks kahest olemasolevast nukleiinhapete tüübist - avas bioloogias uue ajastu, kuna see võimaldas lõpuks mõista, kuidas elusorganismid säilitavad oma elu reguleerimiseks vajalikku teavet ja kuidas nad seda teavet oma järglastele edastavad. . Oleme juba eespool märkinud, et nukleiinhapped koosnevad monomeersetest ühikutest, mida nimetatakse nukleotiidideks. Nukleotiididest ehitatakse üles ülipikad molekulid – polünukleotiidid.
Polünukleotiidide struktuuri mõistmiseks on seetõttu vaja kõigepealt tutvuda sellega, kuidas ehitatud nukleotiidid.
Nukleotiidid. Nukleotiidide struktuur
nukleotiidi molekul koosneb kolmest osast – viie süsinikusisaldusega suhkrust, lämmastikku sisaldavast alusest ja fosforisisaldusest.
Suhkur sisaldub nukleotiidide koostis, sisaldab viit süsinikuaatomit, see tähendab, et see on pentoos. Sõltuvalt nukleotiidis sisalduva pentoosi tüübist on kahte tüüpi nukleiinhappeid - ribonukleiinhapped (RNA), mis sisaldavad riboosi, ja desoksüribonukleiinhapped (DNA), mis sisaldavad desoksüriboosi. Deoksüriboosis asendatakse OH-rühm 2. süsinikuaatomi juures H-aatomiga, see tähendab, et selles on üks hapnikuaatom vähem kui riboosis.
Mõlemas nukleiinhapete tüübid sisaldab nelja erinevat tüüpi aluseid: kaks neist kuuluvad puriinide klassi ja kaks pürimidiinide klassi. Rõngas sisalduv lämmastik annab nendele ühenditele peategelase. Puriinide hulka kuuluvad adeniin (A) ja guaniin (G) ning pürimidiinid tsütosiin (C) ja tümiin (T) või uratsiil (U) (vastavalt DNA-s või RNA-s). Tümiin on keemiliselt väga lähedane uratsiilile (see on 5-metüüluratsiil, st uratsiil, milles 5. süsinikuaatomi juures on metüülrühm). Puriini molekulil on kaks tsüklit, pürimidiini molekulil aga üks.
Vundamendid Nende nime esimene täht on tavaks tähistada: A, G, T, U ja C.

Nukleiinhapped on happed, kuna nende molekul sisaldab fosforhapet.
Joonisel on näha, kuidas suhkur, alus ja fosforhape ühinevad nukleotiidi molekul. Suhkru ja alusega kombineerimine toimub veemolekuli vabanemisel, see tähendab, et see on kondensatsioonireaktsioon. Nukleotiidi moodustamiseks on vaja veel ühte kondensatsioonireaktsiooni - suhkru ja fosforhappe vahel.
Mitmesugused nukleotiidid erinevad üksteisest suhkrute ja nende hulka kuuluvate aluste olemuse poolest.
Nukleotiidide roll organismis ei ole piiratud nukleiinhapete ehitusplokkidena toimimisega; mõned olulised koensüümid on ka nukleotiidid. Need on näiteks adenosiintrifosfaat (ATP), tsükliline adenosiinmonofosfaat (cAMP), koensüüm A, (NAD), nikotii(NADP) ja flaviinadeniini dinukleotiid (FAD).
Nukleiinhapped on looduslikud makromolekulaarsed ühendid (polünukleotiidid), millel on tohutu roll elusorganismide päriliku teabe säilitamisel ja edastamisel.
Nukleiinhapete molekulmass võib varieeruda sadadest tuhandetest kümnete miljarditeni. Need avastati ja eraldati raku tuumadest juba 19. sajandil, kuid nende bioloogiline roll selgus alles 20. sajandi teisel poolel.
Nukleotiidi - nukleiinhapete struktuuriüksuse - koostis sisaldab kolme komponenti:
1) lämmastikalus - pürimidiin või puriin
Pürimidiini alused- pürimidiini derivaadid, mis on osa nukleiinhapetest:uratsiil, tümiin, tsütosiin.
Aluste jaoks, mis sisaldavad rühma –OH, on iseloomulik struktuursete isomeeride liikuv tasakaal, mis on tingitud prootoni ülekandest hapnikust lämmastikule ja vastupidi:
Puriini alused- nukleiinhapete osaks olevad puriini derivaadid: adeniin, guaniin.
Guaniin eksisteerib kahe struktuurse isomeerina:
2) monosahhariid
Riboos ja 2-desoksüriboos Termin "monosahhariidid" viitab viit süsinikuaatomit sisaldavatele monosahhariididele. Need sisalduvad nukleiinhapete koostises tsüklilistes β-vormides:
3) fosforhappe jääk
DNA ja RNA
Sõltuvalt sellest, milline monosahhariid sisaldub polünukleotiidi struktuuriüksuses - riboos või 2-desoksüriboos, eristama
· ribonukleiinhapped(RNA) ja
· desoksüribonukleiinhapped(DNA)
RNA peamine (suhkru-fosfaat) ahel sisaldab jääke riboos ja DNA-s 2-desoksüriboos.
DNA makromolekulide nukleotiidühikud võivad sisaldada adeniin, guaniin, tsütosiin ja tümiin. RNA koostis erineb selle asemel tümiin kohal uratsiil.
DNA molekulmass ulatub kümnete miljonite amu-ni. Need on pikimad teadaolevad makromolekulid. RNA molekulmass on palju väiksem (mitu sada kuni kümneid tuhandeid). DNA-d leidub peamiselt rakkude tuumades, RNA-d - rakkude ribosoomides ja protoplasmas.
Nukleiinhapete struktuuri kirjeldamisel võetakse arvesse makromolekulide erinevaid organiseerituse tasemeid:esmane ja teisejärguline struktuur.
· Esmane struktuur nukleiinhapped on nukleotiidide koostis ja teatud nukleotiidühikute järjestus polümeeri ahelas.
Näiteks:
Lühendatud ühetähelises tähises on see struktuur kirjutatud kujul
...– A – G – C –...
· Under sekundaarne struktuur nukleiinhapped mõistavad polünukleotiidahelate ruumiliselt järjestatud vorme.
DNA sekundaarne struktuurkoosneb kahest paralleelsest hargnemata polünukleotiidahelast, mis on keerdunud ümber ühise telje topeltheeliksiks.
Sellist ruumistruktuuri hoiavad paljud vesiniksidemed, mis on moodustatud spiraali sissepoole suunatud lämmastikualustest.Vesiniksidemed tekivad ühe ahela puriinaluse ja teise ahela pürimidiinaluse vahel. Need alused moodustavad täiendavaid paare (alates lat. komplementum- lisamine).
Vesiniksidemete moodustumine komplementaarsete aluspaaride vahel on tingitud nende ruumilisest vastavusest.
Pürimidiini alus täiendab puriini alust:
Vesiniksidemed teiste aluspaaride vahel ei võimalda neil sobituda kaksikheeliksi struktuuriga. Seega
TÜMIIN (T) täiendab adeniini (A),
TSÜTOSIIN (C) täiendab GUANIINI (G).
Aluse komplementaarsus määrabahela komplementaarsusDNA molekulides.
Polünukleotiidahelate komplementaarsus on DNA põhifunktsiooni – pärilike tunnuste säilitamise ja edasikandumise – keemiliseks aluseks.
DNA võime mitte ainult säilitada, vaid ka kasutada geneetilist teavet, määratakse selle järgmiste omadustega:
DNA molekulid on võimelised replikatsiooniks (kahekordistuma), st. võimaldab sünteesida teisi algsetega identseid DNA molekule, kuna topeltheeliksi ühes ahelas olev aluste järjestus kontrollib nende asukohta teises ahelas.
DNA molekulid suudavad täiesti täpselt ja kindlalt suunata antud liigi organismidele omaste valkude sünteesi.
RNA sekundaarne struktuur
Erinevalt DNA-st koosnevad RNA molekulid ühest polünukleotiidahelast ja neil puudub rangelt määratletud ruumiline kuju (RNA sekundaarne struktuur sõltub nende bioloogilistest funktsioonidest).
RNA peamine roll on otsene osalemine valkude biosünteesis.
On teada kolme tüüpi raku RNA-d, mis erinevad oma asukoha poolest rakus, koostise, suuruse ja omaduste poolest, mis määravad nende spetsiifilise rolli valgu makromolekulide moodustamisel:
informatsioonilised (maatriks) RNA-d edastavad DNA-sse kodeeritud informatsiooni valgu struktuuri kohta raku tuumast ribosoomidesse, kus toimub valgusüntees;
transport-RNA-d koguvad aminohappeid raku tsütoplasmasse ja viivad need ribosoomi; Seda tüüpi RNA molekulid "õpivad" messenger-RNA ahela vastavatest osadest, millised aminohapped peaksid valkude sünteesis osalema;
Ribosomaalsed RNA-d tagavad teatud struktuuriga valkude sünteesi, lugedes teavet informatsioonilisest (maatriks) RNA-st.
on komplekssed monomeerid, millest on kokku pandud heteropolümeeri molekulid. DNA ja RNA. Vabad nukleotiidid osalevad elu signaali- ja energiaprotsessides. DNA nukleotiididel ja RNA nukleotiididel on ühine struktuuriplaan, kuid need erinevad pentoossuhkru struktuuri poolest. DNA nukleotiidid kasutavad suhkru desoksüriboosi, RNA nukleotiidid aga riboosi.
Nukleotiidi struktuur
Iga nukleotiidi võib jagada kolmeks osaks:
1. Süsivesik on viieliikmeline pentoossuhkur (riboos või desoksüriboos).
2. Fosfori jääk (fosfaat) on fosforhappe jääk.
3. Lämmastikalus on ühend, milles on palju lämmastikuaatomeid. Nukleiinhapetes kasutatakse ainult 5 tüüpi lämmastiku aluseid: adeniin, tümiin, guaniin, tsütosiin, uratsiil. DNA-s on 4 tüüpi: adeniin, tümiin, guaniin, tsütosiin. RNA-s on samuti 4 tüüpi: Adeniin, Uratsiil, Guaniin, Tsütosiin.On lihtne näha, et RNA-s on tümiin asendatud Uratsiiliga võrreldes DNA-ga.
Pentoosi (riboos või desoksüriboos) üldine struktuurivalem, mille molekulid moodustavad nukleiinhapete "skeleti":
Kui X asendatakse H-ga (X = H), saadakse desoksüribonukleosiidid; kui X on asendatud OH-ga (X = OH), siis saadakse ribonukleosiidid. Kui asendame R asemel lämmastikaluse (puriin või pürimidiin), saame konkreetse nukleotiidi.
Oluline on pöörata tähelepanu süsinikuaatomite positsioonidele pentoosis, mis on tähistatud kui 3" ja 5". Süsinikuaatomite nummerdamine algab ülaosas olevast hapnikuaatomist ja läheb päripäeva. Saadakse viimane süsinikuaatom (5"), mis asub väljaspool pentoositsüklit ja moodustab, võib öelda, et pentoosi "saba". Seega saab nukleotiidide ahela ülesehitamisel ensüüm siduda ainult uue nukleotiidi süsinikule 3 "ja mitte mingile muule . Seetõttu ei saa nukleotiidahela 5-tollist otsa kunagi jätkata; ainult 3-tollist otsa saab pikendada.



Võrrelge RNA nukleotiidi DNA nukleotiidiga.
Proovige välja selgitada, mis nukleotiid see on selles esituses:

ATP - vaba nukleotiid
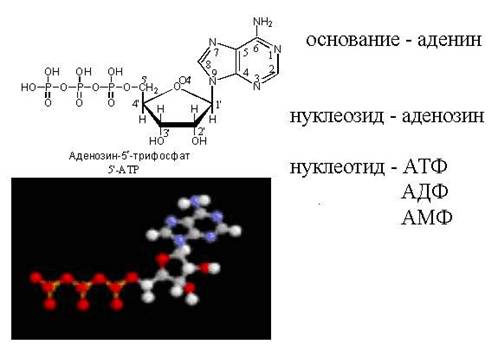
cAMP – "loopback" ATP molekul

Nukleotiidi struktuuri skeem

Pange tähele, et aktiveeritud nukleotiidil, mis on võimeline üles ehitama DNA või RNA ahelat, on "trifosfaadi saba". Just selle "energiaga küllastunud" sabaga saab ta liituda juba olemasoleva kasvava nukleiinhappe ahelaga. Fosfaadi saba asub süsinikul 5, nii et süsiniku asend on juba fosfaatide poolt hõivatud ja see on mõeldud kinnitamiseks. Mille külge seda kinnitada? Ainult süsinikule positsioonis 3". Kui see on kinnitatud, muutub see nukleotiid ise sihtmärgiks järgmise nukleotiidi kinnitumisel. "Vastuvõttev pool" annab süsiniku positsioonis 3" ja "sissetulev pool" klammerdub selle külge. fosfaadi saba, mis asub positsioonis 5". Üldiselt kasvab kett 3" küljelt.
DNA nukleotiidahela pikendamine
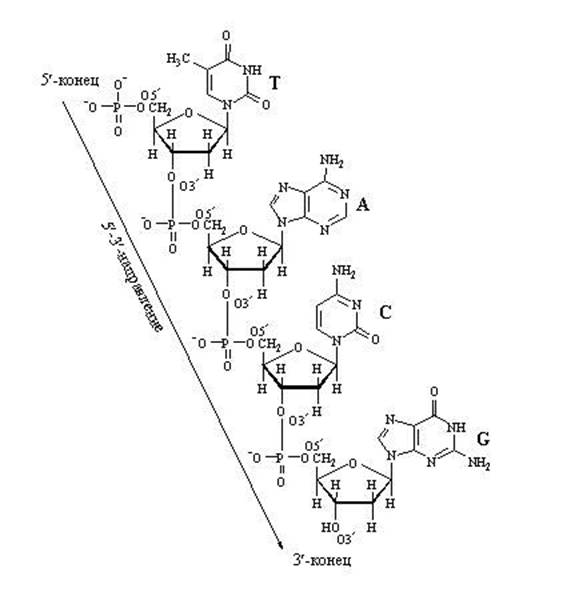
Nukleotiidide vaheliste "pikisuunaliste" sidemete tõttu võib ahela kasv kulgeda ainult ühes suunas: 5" ⇒ kuni 3", sest Uue nukleotiidi saab lisada ainult ahela 3'-otsa, mitte 5'-otsa.
Nukleotiidide paarid, mis on ühendatud nende lämmastikualuste "ristsete" komplementaarsete sidemetega
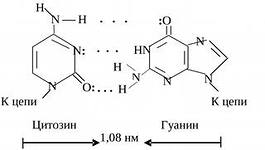
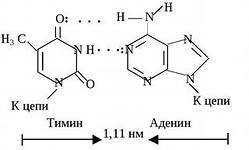
DNA kaksikheeliksi osa

Leidke kahe DNA ahela antiparallelsuse tunnused.
Leidke topelt- ja kolmiksidemetega nukleotiidide paarid.