Millised elundid moodustuvad idukihtidest. idukihi teooria. Idukihtide derivaadid. Idukihtide teooria kujunemise ajalugu
Idukihid on embrüoloogia põhitermin. Need määravad loote keha kihid juba varajases staadiumis.Enamasti on need kihid oma olemuselt epiteelsed.
Idukihid jagunevad tavaliselt kolme tüüpi:
Ektoderm - välimine kiht, mida nimetatakse ka epiblastiks või nahatundlikuks kihiks;
Endoderm on rakkude sisemine kiht. Seda võib nimetada ka hüpoblastiks või entero-näärmeleheks;
Keskmine kiht (mesoderm või mesoblast).
Idulehed (olenevalt nende asukohast iseloomustavad neid teatud rakutunnused. Seega koosneb embrüo välimine kiht heledatest ja kõrgetest rakkudest, mis on ehituselt sarnased silindrilise epiteeliga. Siseleht koosneb enamikul juhtudel suurtest rakkudest mis on täidetud spetsiifiliste munakollaste plaatidega. Need on lameda välimusega, mis muudab need sarnaseks
Mesoderm koosneb esimeses etapis spindlikujulistest ja tähtkujulistest rakkudest. Hiljem moodustavad nad epiteelikihi. Pean ütlema, et paljud teadlased usuvad, et mesoderm on keskmised idukihid, mis ei ole iseseisev rakukiht.
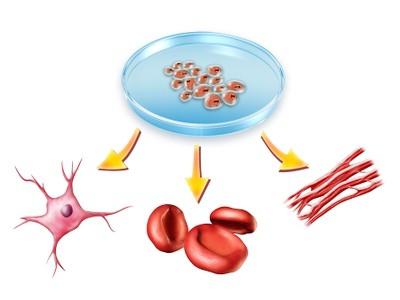
Idukihid on algul õõnsa moodustisena, mida nimetatakse blastodermaalseks vesiikuliks. Ühele selle poolustele koguneb rühm rakke, mida nimetatakse rakumassiks. See tekitab primaarse soole (endodermi).
Olgu öeldud, et embrüonaalsetest lehtedest moodustuvad erinevad elundid. Seega tekib ektodermist närvisüsteem, endodermist pärineb seedetoru ning mesodermist skelett ja lihased.
Samuti tuleb märkida, et embrüogeneesi käigus moodustuvad spetsiaalsed embrüonaalsed membraanid. Need on ajutised, ei osale elundite moodustamises ja eksisteerivad ainult embrüonaalse arengu ajal. Igal klassil on nende kestade moodustamisel ja struktuuril teatud tunnused.

Embrüoloogia arenguga hakkasid nad määrama embrüote sarnasust, mida kirjeldas esmakordselt K.M. Baer 1828. aastal. Veidi hiljem tuvastas Charles Darwin kõigi organismide embrüote sarnasuse peamise põhjuse - nende ühise päritolu. Severov aga väitis, et embrüote ühised tunnused on seotud evolutsiooniga, mis enamikul juhtudel kulgeb anabolismi kaudu.
Erinevate klasside ja loomaliikide embrüote peamiste arengufaaside võrdlemisel leiti teatud tunnused, mis võimaldasid sõnastada embrüonaalse sarnasuse seaduse. Selle seaduse põhisätted seisnesid selles, et sama tüüpi organismide embrüod nende arengu algstaadiumis on väga sarnased. Seejärel iseloomustavad embrüot üha rohkem individuaalseid tunnuseid, mis viitavad tema kuulumisele vastavasse perekonda ja liiki. Samal ajal eraldatakse sama tüüpi esindajate embrüod üksteisest üha enam ja nende esmast sarnasust enam ei tuvastata.
idukihid(lat. embrüonaalne lehestik), idukihid, mitmerakuliste loomade embrüo keha kihid, mis moodustuvad gastrulatsiooni käigus ja annavad alguse erinevatele elunditele ja kudedele. Enamikus organismides moodustub kolm idukihti: välimine on ektoderm, sisemine endoderm ja keskmine mesoderm.
Ektodermi derivaadid täidavad peamiselt integumentaalseid ja sensoorseid funktsioone, endodermi derivaadid - toitumis- ja hingamisfunktsioonid ning mesodermi derivaadid - embrüo osade vahelised ühendused, motoorsed, tugi- ja troofilised funktsioonid.
Erinevate selgroogsete klasside esindajatel on sama idukiht samade omadustega, s.t. idukihid on homoloogsed moodustised ja nende olemasolu kinnitab loomamaailma päritolu ühtsuse positsiooni. Idukihid moodustuvad kõigi selgroogsete suuremate klasside embrüote, s.o. on universaalselt levinud.
Idukiht on rakkude kiht, mis hõivab teatud positsiooni. Kuid seda ei saa käsitleda ainult topograafiliste positsioonide järgi. Idukiht on rakkude kogum, millel on teatud arengutendentsid. Selgelt määratletud, kuigi üsna lai arengupotentsiaalide vahemik määratakse (määratakse) lõpuks gastrulatsiooni lõpuks. Seega areneb iga idukiht kindlas suunas, osaleb teatud elundite alge tekkes. Kogu loomariigis pärinevad üksikud elundid ja koed samast idukihist. Ektodermist moodustub neuraaltoru ja siseepiteel, endodermist - sooleepiteel, mesodermist - lihas- ja sidekude, neerude epiteel, sugunäärmed ja seroossed õõnsused. Mesodermist ja kraniaalsest osast väljutatakse ektodermi rakud, mis täidavad lehtedevahelise ruumi ja moodustavad mesenhüümi. Mesenhümaalsed rakud moodustavad süntsütiumi: need on omavahel ühendatud tsütoplasmaatiliste protsesside kaudu. Mesenhüüm moodustab sidekoe. Iga üksik idukiht ei ole autonoomne moodustis, see on osa tervikust. Idukihid suudavad eristuda ainult üksteisega suheldes ja embrüo kui terviku integreerivate mõjude mõju all. Sellise vastastikuse mõju ja vastastikuse mõjutamise hea näide on katsed kahepaikse gastrula varajase kohta, mille kohaselt saab ekto-, ento- ja mesodermi rakumaterjali sundida radikaalselt muutma oma arenguteed, osalema elundite moodustamises. mis on sellele infolehele täiesti ebaloomulikud. See viitab sellele, et gastrulatsiooni alguses ei ole iga idukihi rakumaterjali saatus rangelt võttes veel ette määratud. Iga lehe areng ja diferentseerumine, nende organogeneetiline spetsiifilisus on tingitud kogu embrüo osade vastastikusest mõjust ja on võimalik ainult normaalse integratsiooni korral.
62. Histo- ja organogenees. Neurulatsiooni protsess. Teljeorganid ja nende teke. mesodermi diferentseerumine. Selgroogsete embrüote tuletisorganid.
Histogenees(muu kreeka keelest ἱστός - kude + γένεσις - teke, areng) - protsesside kogum, mis viib kudede moodustumiseni ja taastumiseni isendi arengu (ontogeneesi) käigus. Teatud tüüpi koe moodustumisel osaleb üks või teine idukiht. Näiteks lihaskude areneb mesodermist, närvikude ektodermist jne. Mõnel juhul võivad sama tüüpi kuded olla erineva päritoluga, näiteks naha epiteel on ektodermaalne ja absorbent sooleepiteel on endodermaalne. päritolu.
Organogenees- embrüonaalse isendi arengu viimane etapp, millele eelneb viljastamine, purustamine, blastulatsioon ja gastrulatsioon.
Organogeneesis, neurulatsioonis, histogeneesis ja organogenees.
Neurulatsiooni käigus moodustub neurul, millesse asetatakse mesoderm, mis koosneb kolmest idukihist (mesodermi kolmas kiht jaguneb segmenteeritud paarisstruktuurideks - somiitideks) ja aksiaalsest elundite kompleksist - neuraaltorust, akordist. ja soolestikku. Elundite aksiaalse kompleksi rakud mõjutavad üksteist vastastikku. Seda vastastikust mõju nimetatakse embrüonaalseks induktsiooniks.
Histogeneesi protsessis moodustuvad kehakuded. Ektodermist moodustub närvikude ja naha epidermis koos nahanäärmetega, millest järgnevalt arenevad närvisüsteem, meeleelundid ja epidermis. Endodermist moodustub notokord ja epiteelkude, millest järgnevalt moodustuvad limaskestad, kopsud, kapillaarid ja näärmed (välja arvatud suguelundite ja naha omad). Mesoderm toodab lihas- ja sidekude. ODS, veri, süda, neerud ja sugunäärmed moodustuvad lihaskoest.
Neurulatsioon- neuraalplaadi moodustumine ja selle sulgemine neuraaltorusse akordide embrüonaalse arengu protsessis.
Neurulatsioon on ontogeneesi üks peamisi etappe. Neurulatsiooni staadiumis embrüot nimetatakse neurulaks.
Neuraaltoru arengut ees-tagasuunas juhivad spetsiaalsed ained - morfogeenid (need määravad, milline otstest saab aju) ja selle kohta käiv geneetiline teave on põimitud nn homöootilistesse või homöootilistesse geenidesse.
Näiteks morfogeen retinoehape on selle kontsentratsiooni suurenemisega võimeline muutma ühte tüüpi rombomeere (aju tagumise osa neuraaltoru segmendid) teiseks.
Lantsettide neurulatsioon on ektodermist pärit ribide kasv üle rakukihi, millest saab närviplaat.
Neurulatsioon kihilises epiteelis - mõlema kihi rakud laskuvad segatuna ektodermi alla ja lahknevad tsentrifugaalselt, moodustades neuraaltoru.
Neurulatsioon ühekihilises epiteelis:
Skisokoeloosne tüüp (teleostides) - sarnane kihilise epiteeli neurulatsiooniga, välja arvatud see, et ühe kihi rakud laskuvad alla.
Lindudel ja imetajatel tungib neuraalplaat sissepoole ja sulgub neuraaltorusse.
Lindudel ja imetajatel neurulatsiooni ajal neuraalplaadi väljaulatuvad osad nn närvivoldid, on suletud kogu neuraaltoru pikkuses ebaühtlaselt.
Tavaliselt sulgub esmalt neuraaltoru keskosa ja seejärel läheb sulgur mõlemasse otsa, jättes selle tulemusena lahti kaks osa – eesmine ja tagumine neuropoor.
Inimestel on neuraaltoru sulgemine keerulisem. Seljaosa sulgub esiteks rindkere nimmepiirkonnast, teine - ala otsaesist kroonini, kolmas - eesmine, läheb ühes suunas, neurokraniumini, neljas - piirkond, mis asub rindkere tagant. pea emakakaela lõpuni, viimane, viies - sakraalne osa, läheb samuti ühte suunda, koksiuksist eemale.
Kui teine sektsioon ei ole suletud, leitakse surmav kaasasündinud defekt - anentsefaalia. Lootel aju ei arene.
Kui viiendat sektsiooni ei suleta, leitakse kaasasündinud defekt, mida saab parandada – spina bifida ehk Spinabifida. Sõltuvalt raskusastmest jaguneb spina bifida mitmeks alatüübiks.
Neurulatsiooni käigus moodustub neuraaltoru.
Ristlõikes, kohe pärast moodustamist, saab selles eristada kolme kihti, seestpoolt väljapoole:
Ependüüm – algelisi rakke sisaldav pseudokihiline kiht.
Vahevöötsoon sisaldab rändavaid, vohavaid rakke, mis väljuvad ependüümikihist.
Välimine marginaalne tsoon on kiht, kus moodustuvad närvikiud.
Neid on 4 aksiaalne keha: notokord, neuraaltoru, sooletoru ja mesoderm.
Sõltumata loomaliigist muundatakse need rakud, mis migreeruvad läbi blastopoori dorsaalse huule piirkonna, seejärel notokordiks ja läbi blastopoori külgmiste (külgmiste) huulte piirkonna kolmandaks idukihiks - mesodermiks. Kõrgematel akordidel (linnud ja imetajad) idukilbi rakkude immigratsiooni tõttu blastopoor gastrulatsiooni ajal ei moodustu. Rakud, mis migreerusid läbi blastopoori dorsaalse huule, moodustavad kõõlu, tiheda raku nööri, mis paikneb piki embrüo keskjoont ektodermi ja endodermi vahel. Selle mõjul hakkab välimises idukihis tekkima neuraaltoru ja alles viimasena moodustab endoderm sooletoru.
Mesodermi diferentseerumine (lat. differents. different) algab 3. arengunädala lõpus. Mesenhüüm tekib mesodermist.
Mesodermi dorsaalne osa, mis paikneb kõõlu külgedel, jaguneb keha segmentideks – somiitideks, millest arenevad luud ja kõhred, vöötskeletilihased ja nahk (joon. 134).
Mesodermi ventraalsest segmenteerimata osast - planchnotoomiga - moodustuvad kaks plaati: splanchnopleura ja somatopleura, millest areneb seroossete membraanide mesoteel ja nendevaheline ruum muutub kehaõõnsusteks, seedetoruks, vererakud, silelihaskoe, vere- ja lümfisooned, sidekude, südame vöötlihaskoe, neerupealise koor ja epiteel sugunäärmed.
Idukihtide derivaadid. Ektodermist moodustuvad väliskesta, kesknärvisüsteem ja seedekanali lõpposa. Endodermist moodustub notokord, seedetoru keskmine osa ja hingamissüsteem. Mesodermist moodustuvad luu- ja lihaskonna, kardiovaskulaarsed ja urogenitaalsüsteemid.
Looge vastavus inimkeha struktuuri ja idukihi vahel, millest see moodustati.
Kirjutage vastuseks numbrid üles, korraldades need tähtedele vastavas järjekorras:
| A | B | AT | G | D |
Selgitus.
Olulisemad ektodermaalsed derivaadid on neuraaltoru, närvihari ja kõik neist moodustunud närvirakud. Ektodermaalsetest angaaridest arenevad ka meeleorganid, mis edastavad närvisüsteemile infot nägemis-, heli-, haistmis- ja muude stiimulite kohta. Näiteks silma võrkkest moodustub aju väljakasvuna ja on seetõttu neuraaltoru derivaat, samas kui haistmisrakud eristuvad otse ninaõõne ektodermaalsest epiteelist. Valu retseptorid on ektodermaalset päritolu.
Ektoderm: valuretseptorid, juuksepiir, küüneplaadid. Mesoderm: lümf ja veri, rasvkude.
Vastus: 11221.
Vastus: 11221
Allikas: Bioloogia ühtne riigieksam 30.05.2013. põhilaine. Siber. 2. variant.
Sadi 11.06.2017 13:49
Selle ülesande vastuses on kirjas, et kopsud moodustuvad mesodermist ja ülesandes 8 nr 13837 on öeldud, et endodermist.
Natalja Evgenievna Bashtannik
Pange tähele, et kopsude epiteel on endoderm.
Konkreetse elundi rudiment moodustub esialgu konkreetsest idukihist, kuid seejärel muutub elund keerukamaks ja selle moodustumisel osaleb selle tulemusena kaks-kolm idukihti.
Kops ei ole ainult epiteel, see on ka bronhioolid ja sidekiled ... see kõik moodustub mesenhüümist ja kahjuks ei arvestata seda teadmist juba ühtsel riigieksamil :(
Arenevate bronhide vaheline ruum on täidetud vahepealse mesenhüümiga. Mesenhüüm, mis on lahtine kude, mis katab tihedalt arenevaid endodermaalseid torukujulisi moodustisi, hakkab diferentseeruma kolmandal kuul kopsujuures. Siit jätkub diferentseerumine perifeerses suunas eraldi bronhide harudega. Esiteks ilmuvad mõlema peamise bronhi kõhrerõngad ja järk-järgult diferentseeruvad ülejäänud bronhide kõhreplaadid. Ligikaudu samal ajal moodustuvad lihasrakud ja sidekoe esimesed kollageenkiud. Mesodermaalsest materjalist tekib kopsukile interlobulaarne ja intersegmentaalne vaheseina mesenhüüm ning subseroosne sidekude. Elastsed kiud hakkavad ilmuma neljandal kuul. Nende peamine areng, nagu ka kõhreplaatide areng bronhide seintes, toimub siiski alles emakasisese arengu teisel poolel.
"MORDOVA RIIKLIKÜLIKOOL, mille nimi on A.I. N. P. OGAREVA»
bioloogia osakond
Geneetika osakond
teemal: idukihid
Lõpetanud: 3. kursuse üliõpilane
eriala "bioloogia"
Sissejuhatus
1. Idukihtide struktuur
2. Idukihtide teooria kujunemislugu
3. Idukihtide teke
4. Idukihtide päritolu ja evolutsiooniline tähtsus
5. Idukihtide teooria sätted ja vastuväited sellele teooriale
Järeldus
Kirjandus
Sissejuhatus
Lisaks võimalusele tõlgendada idukihte nende fülogeneetilise tähtsuse seisukohalt, on oluline kindlaks teha nende roll isendi arengus. Idukihid on esimesed organiseeritud rakurühmad embrüos, mis eristuvad üksteisest selgelt oma tunnuste ja suhete poolest. Asjaolu, et need suhted on kõigi selgroogsete embrüote puhul põhimõtteliselt samad, viitab tugevalt selle tohutu loomarühma erinevate liikmete ühisele päritolule ja sarnasele pärilikkusele.
Võib arvata, et neis idukihtides hakkavad esmakordselt tekkima erinevate klasside erinevused kehaehituse üldplaani kohal, mis on iseloomulikud kõigile selgroogsetele.
Idukihtide moodustumine lõpetab perioodi, mil peamiseks arenguprotsessiks on vaid rakkude arvu suurenemine ning algab rakkude diferentseerumise ja spetsialiseerumise periood. Diferentseerumine toimub idukihtides enne, kui me näeme selle märke ühegi meie mikroskoopilise meetodiga. Täiesti ühtlase välimusega lehes tekivad pidevalt lokaliseeritud rakurühmad, millel on erinev edasiarengu potentsiaal.
Idukihist tekivad mitmesugused struktuurid. Samal ajal ei ole idukihis märgatavad nähtavad muutused, mille tõttu need tekivad. Hiljutised eksperimentaalsed uuringud näitavad, kui varakult eelneb see nähtamatu diferentseerumine rakurühmade nähtavale morfoloogilisele lokaliseerimisele, mida tunneme kergesti ära kui lõpliku organi alge.
1. Idukihtide struktuur
Idukihid koosnevad rakulistest materjalidest, mida kasutatakse erinevate elundite ja kudede arendamiseks. Oma struktuurilt erinevad erinevate idukihtide rakud üksteisest; endodermi rakud on alati suuremad ja vähem korrapärased kui ektodermaalsed rakud. Endodermi eristavad tulevase järjehoidja omadused, millel on troofiline tähendus. Ektoderm jääb pinnale ja on esialgu kaitsva väärtusega. Erinevalt endodermist koosneb see korrapäraselt paigutatud ühtlasema kujuga rakkudest. Gastrulatsioon toob kaasa märgatava erinevuse välimise ja sisemise kihi vahel ning idumaterjal muutub heterogeenseks. Protsessi, mis viib algselt homogeenses materjalis erinevuste ilmnemiseni, nimetatakse diferentseerumiseks.
Põhiorganisaatorid ehk induktiivpoolid mängivad olulist rolli rakulise materjali eristamisel. Induktorid on kemikaalid, mis vabanevad rakurühmadest ja mõjutavad teisi rakurühmi, muutes nende arenguteed. Idukihtide diferentseerumise tulemusena moodustuvad mitmesugused elundid ja koed. Nende protsesside uurimisel erinevatel loomadel leiti, et kõigi hulkraksete organismide iga idukihi saatus on reeglina sama.
Nii arenevad ektodermist välja naha epiteel, nahanäärmed, paljud sarve derivaadid, närvisüsteem ja meeleelundid. Endodermist moodustub kõigil loomadel sooletrakti keskosa epiteel, maks ja seedenäärmed. Kordaatides moodustub ka hingamisteede epiteel. Mesodermist arenevad veri ja lümf, lihas-, side-, kõhre- ja luukoed, neeruepiteel, sekundaarse kehaõõne sein, osa reproduktiivsüsteemi kudesid.
2. Idukihtide teooria kujunemislugu
Idukihi teooria on üks suurimaid võrdleva embrüoloogia üldistusi 19. sajandil. Idukihte kirjeldas esmakordselt X. Pander (1817), kes avastas, et kana embrüo koosneb teatud arenguetappidel kolmest õhukesest kilest või kihist, mille rakuline olemus polnud veel teada. Pander nimetas välimist lehte seroosseks, sügavaimat - limaskestaks ja vahepealset - vereks. Neid tähelepanekuid kinnitas K. Baer (1828, 1837), kes leidis idukihte ka mõnel teisel loomal (kalad, konnad, kilpkonnad). Baer eristas kahte primaarset kihti - loomset ja vegetatiivset, mis jagunevad seejärel taas sekundaarseteks idukihtideks: loomne kiht annab naha ja lihase ning vegetatiivne - veresoonte ja limaskestade. Kaasaegse terminoloogia järgi vastab nahaleht ektodermile, limaskest endodermile ning lihas- ja vaskulaarne leht mesodermi parietaalsele ja vistseraalsele lehele. Baeri viga seisnes ainult selles, et ta kirjeldas nende kahe mesodermaalse kihi päritolu selgroogsetes erinevatest allikatest. Terminid "ektoderm" ja "endoderm" laenasid embrüoloogid zooloogiast (nii nimetati täiskasvanud cnidarlaste keha moodustavaid epiteelikihte veelgi varem). Kana embrüo idukihtide rakulise struktuuri määras Remak 1855. aastal.
Algselt arvati, et idukihid tekivad alles selgroogsete arengu käigus. Peaaegu kõigi selgrootute klasside arengut uurinud A. O. Kovalevski ja I. I. Mechnikovi tööde järel sai aga selgeks, et idukihid esinevad ühel või teisel kujul kõigil hulkraksete loomade puhul. A. O. Kovalevsky (1871) kirjutas artiklis “Usside ja lülijalgsete embrüoloogilised uuringud” lõpuosas: “Kui nüüd võrrelda meie poolt kirjeldatud usside arengut teiste loomade arenguga, siis idukihtide analoogia selgroogsete omad on meie jaoks eriti silmatorkavad, kuni üksikute detailideni välja; samad kaks peamist lehte, millel on suur roll usside arengus, on ka selgroogsetel; nagu mõnel, nii ka teistel, ilmub keskmine leht alles hiljem. Lehtede saatused ja elundite munemine langevad äärmiselt kokku, kuni üksikute protsessideni välja.
I. I. Mechnikov avastas mõnel loomal tugevalt muutunud arenguga idukihid ja tõstatas esimest korda küsimuse gastrulatsiooniprotsesside evolutsioonist.
3. Idukihtide teke
Idukihid tekivad loomadel ja inimestel protsessis, mida nimetatakse gastrulatsiooniks.
Loomadest eristatakse kahe- ja kolmekihilisi taksoneid. Alates lameussidest on loomadel 3 idukihti: ektoderm (välimine), endoderm (sisemine) ja mesoderm (keskmine). Mesodermi leidub ainult kolmekihilistel loomadel, ektodermi ja endodermi aga kahekihilistel (käsnad, sammalloomad, koelenteraadid) ja kolmekihilistel loomadel.
Närvisüsteem, nahk, nahanäärmed, naha derivaadid nagu suled, juuksed, küüned, küünised, soomused, samuti seedetoru eesmise ja tagumise osa epiteel ning vistseraalse luustiku luud arenevad. ektoderm ontogeneesis.
Soole limaskesta moodustub endodermist; endoderm toidab embrüot; sellest idukihist arenevad hingamiselundid, seedesüsteemi limaskestad ja seedenäärmed (maks jne).
Mesodermist moodustuvad vereringe-, eritus- ja reproduktiivsüsteemi organid, tsöloomi ja siseorganite seroosmembraanid, samuti tugiskeleti ja lihaste luud.
Kaasaegsed meetodid embrüonaalse protsessi uurimisel on võimaldanud tuvastada, et idukihtidel ei ole primitiivse elundi tähendust ega kordu ühtegi fülogeneetilise arengu etappi. Neid tuleks käsitleda kui materjali teatud tulevaste elundite kompleksist, mis on samal arengutasemel ja on morfoloogiliselt sarnased. Idukihtide moodustumise protsess tähistab teatud elundite arenguetappi, mille läbib valdav enamus loomi.
Tavaliselt koosneb iga elund erinevatest idukihtidest pärinevatest kudedest, kuid me liigitame elundi ühe või teise lehe derivaadiks, olenevalt sellest, millest areneb tema põhialg. Seega koosneb selgroogsete kesksoole sein endodermaalsest epiteelist ja mesodermilistest silelihastest ning sidekoe kihist. Aga kuna kesksoole esimene rudiment moodustub endodermist, millele hiljem kinnituvad mesodermaalsed elemendid ning seedefunktsiooni täidab endodermaalne epiteel, siis peetakse kesksoole endodermaalseks organiks.
Idukihtide olemasolu, mis on sarnaselt seotud kõigi Metazoa kehaehitusega, võimaldas võrrelda süstemaatiliselt kaugete loomarühmade arengut. Praegusel ajal on lihtsalt võimatu kirjeldada ühegi looma arengut ilma idukihte mainimata.
4. Idukihtide päritolu ja evolutsiooniline tähtsus
Tekib küsimus, mis on idukihtide päritolu ja evolutsiooniline tähtsus. E. Haeckeli (1874) järgi korduvad primaarsed idukihid (ekto- ja endoderm) arengus (kokkuvõtvad) metazoa hüpoteetilise ühise esivanema - Gastrea - esmased organid (nahk ja sooled). Sellest järeldub, et kõigi loomade idukihid on homoloogsed. I. I. Mechnikov (1886) omistas idukihtidele ka rekapitulatsiooni tähenduse, kuid ta esindas metazoa ühist esivanemat Phagocytella kujul. Mechnikovi sõnul esindab kinoblasti arengu käigus ektoderm ja kõik kinoblastist evolutsiooni käigus tekkinud elundid on isendi arengu käigus ektodermaalset päritolu. Fagotsütoblasti areng toimus kahes suunas. Koelenteraatides epiteliseerub see täielikult ja muutus maoõõne limaskestaks; individuaalses arengus esindab seda endoderm. Kolmekihilistel loomadel muutus ainult fagotsütoblasti keskosa soolestikuks ja seda esindab ontogeneesis endoderm, perifeerne osa aga tekitas sisekeskkonna kudesid ja seda esindab ontogeneesis mesoderm.
5. Idukihtide teooria sätted ja vastuväited sellele teooriale
Seega XIX sajandi lõpuks. välja on kujunenud klassikaline idukihtide teooria, mille sisuks on järgmised sätted:
1. Kõigi hulkraksete loomade ontogeneesis moodustub kaks-kolm idukihti, millest arenevad kõik elundid.
2. Idukihte iseloomustab teatud asend embrüo kehas (topograafia) ja neid nimetatakse vastavalt ekto-, ento- ja mesodermiks.
3. Idukihid on spetsiifilised, see tähendab, et igaüks neist tekitab rangelt määratletud primordiaid, mis on kõigil loomadel ühesugused.
4. Idukihid koondavad ontogeneesis kõigi metazoade ühise esivanema esmased organid ja on seetõttu homoloogsed.
5. Elundi ontogeneetiline areng ühest või teisest idukihist näitab tema evolutsioonilist päritolu esivanema vastavast esmasest elundist.
Tänaseks on kogunenud palju fakte, mis esmapilgul klassikalise idukihtide teooria raamidesse ei mahu. Seetõttu hakkasid ilmuma väited, et see teooria on aegunud, kriisis ja vajab ülevaatamist. Kõik need kriitikad põhinevad liiga formaalsel evolutsioonivastasel arusaamal idukihtidest. Vaatleme mõningaid olulisimaid vastuväiteid idukihtide teooriale.
1. Asjaolu, et mesoderm võib pärineda nii ektodermist kui ka endodermist, on olnud paljude erimeelsuste objektiks ja see seab kahtluse alla selle ühtsuses idukihina. Paljud autorid peavad vajalikuks eristada mesoblasti (entomesoderm) ja mesenhüümi (ektomesoderm). Kuid erinevused mesodermi nende osade vahel ei ole nii olulised, kui esmapilgul tundub. Spiraalse fragmentatsiooniga vormide puhul pärineb mesenhüüm 2. ja 3. kvarteti mikromeeridest ning mesoblast kuulub 4. nelikusse: kõik need rakud paiknevad piki blastopoori servi, st ekto- ja kvarteti piiritsoonis. endoderm. Mesenhümaalsete elementide migratsioon blastokoeli on osa gastrulatsioonist. Samuti võib oletada, et fagotsütoblasti, mille perifeerset osa esindab mesoderm, evolutsiooniline moodustumine oli pikk protsess ning selle täienemine kinoblasti toimel jätkus väga kaua, mis kajastub ontogeneesis.
2. Mõnel loomal on idukihid esitatud väga keerulisel kujul. Näiteks putukatel ja lindudel täheldatakse nn kahefaasilist või isegi mitmefaasilist gastrulatsiooni, mis justkui laguneb mitmeks iseseisvaks aktiks.Tihtipeale algab organogenees juba enne idukihtide moodustumist, elundite alged on isoleeritud. Idukihid ei ole selgelt väljendatud. Kuid seda olukorda saab kergesti seletada arengu käigus toimunud teisejärgulise muutuse tulemusega. Ei tohi unustada, et kõik ontogeneetilised protsessid alluvad evolutsioonile samal määral kui täiskasvanud loomade organid. Isegi Cnidaria perekonnas on gastrulatsioon läbi teinud märkimisväärse evolutsiooni, mistõttu pole üllatav, et kõrgematel loomadel, kes on Metazoa päritolust kaugel, on gastrulatsiooniprotsessid läbinud nii põhjalikud sekundaarsed muutused. Pigem tuleks imestada, et me eristame neis veel idukihte, olgugi et modifitseeritud kujul.
3. Rangelt määratud lõhustumise korral (nematoodidel, anneliididel, molluskitel, astsiididel) esindavad üksikud blastomeerid või blastomeeride rühmad juba teatud elundite alge. Nii eristatakse rõngasussil Arenicola 64 blastomeeri staadiumis loomapoolusel 4 rakust koosnev nn rosett, mis on tundliku sultani alge, ja ekvatoriaalvööndis on 4. rakurühmad, igas 4 - trohoblastid, millest areneb prototroh. Vegetatiivsel poolusel on 7 suurt munakollaserikast rakku - soolestiku alge, millega tulevasest seljaküljest külgneb rakk, mis tekitab mesodermaalseid teloblaste. Jääb mulje, et hiljem tekkinud idukihtidel ei ole iseseisvat tähendust, vaid need on vaid ajutine kooslus juba olemasolevatest heterogeensetest algetest.
See ürgsete idukihtide seos ei ole aga juhuslik, vaid on ajalooliselt tingitud. Niisiis hõlmab ektodermi koostis ainult nende organite algeid, mis sellest arenevad ja millel on mittedeterministlik killustatus (nahk, meeleelundid jne). Lisaks on blastomeeride varajane määramine ka sekundaarsete muutuste tagajärg arengu käigus - see on kohanemine, mis võimaldab embrüol kiiresti muutuda vastseks, mis koosneb veel mõnest rakust, kuid on juba võimeline iseseisvalt tegema kõiki. elutähtsad funktsioonid (välja arvatud muidugi seksuaalne).
4. Idukihi teooria kriitikud osutavad tavaliselt mitmesuguste erandite olemasolule, mille hulka kuuluvad käsnade idukihtide moonutamine, selgelt väljendunud kihtide puudumine paljudel lameussidel, endodermi puudumine enamikul sammalloomadel jne. kõik need konkreetsed näited koos nende loomade arengu üksikasjaliku kirjeldusega. Märgime ainult, et kõigi konkreetsete kõrvalekallete esinemist üldreeglist saab evolutsioonilisest vaatenurgast täielikult mõista ja nende põhjused on enamikul juhtudel selged. Lisaks täheldatakse neid kõrvalekaldeid tavaliselt üsna madala organiseeritusega loomadel, samas kui kõrgematel loomadel (lülijalgsed, selgroogsed) järgitakse rangelt idukihtide spetsiifilisust. See viitab sellele, et alumiste metazoa idukihid on väga labiilsed, samas kui nende spetsiifilisus ilmnes hiljem ja areneb evolutsiooni käigus.
5. Mittesugulisel paljunemisel, mitmesugustel taastamisprotsessidel ja eksperimentaalsel sekkumisel arengu käigus täheldatakse sageli idukihtide spetsiifilisuse põhimõtte rikkumist. Niisiis, sammalloomade ja mõnede astsiidlaste tärkamise ajal ei sisaldu endodermaalse iseloomuga kuded neerude koostises ja soolestik areneb ektodermist. Nemertina Lineus lacteus’e puhul saab ära lõigata väikese suueelse kehaosa, mis samuti ei sisalda endodermaalseid organeid ja sellest fragmendist areneb terve loom.
Nende nähtuste olemuse mõistmiseks on vaja meeles pidada, millel põhineb idukihtide eripära. Embrüogeneesis arenevad igast lehest need elundid, mis on ajalooliselt eraldunud vastava rakukihi koostisest, st lehtede spetsiifilisus põhineb rekapitulatsiooni fenomenil. Kokkuvõte ise (nagu on näidanud I. I. Shmalgauzen) on suuresti tingitud asjaolust, et embrüo osade vahel on teatud ajalooliselt väljakujunenud morfogeneetilised korrelatsioonid. Kuid taastumisprotsesside ja mittesugulise paljunemise käigus ei toimu areng mitte gastrula, vaid täiskasvanud looma kudede põhjal, mille vahel on muid füsioloogilisi seoseid. Idukihid on eranditult embrüonaalsed moodustised ja sellisena puuduvad täiskasvanud loomadel. Seetõttu kaotab idukihtide spetsiifilisus oma tähtsuse.
Sellele võib lisada, et mittesugulise paljunemise võime ja kudede laiemad morfogeneetilised võimed on iseloomulikud vaid loomadele, kes ei ole jõudnud väga kõrgele evolutsioonilisele tasemele, mis viitab täiskasvanud looma idukihtide ja kudede progresseeruvale spetsiifilisusele.
Kaasaegset vaatenurka idukihtidele väljendab hästi järgnev tsitaat V. N. Beklemiševi teosest “Selgrootute võrdlev anatoomia”: “... kinoblast ja fagotsütoblast on keha põhikihid ja looma otsesed elundid alles aastal. koelenteraatide ja käsnade vastsed ning kõige lihtsamini paigutatud hüdroidide vastsed, nagu Protohydra. Kõigis teistes Enterozoades lagunevad esmased kihid funktsioonide koondumise ja elundite integratsiooni tõttu mitmeteks derivaatideks, mis on omavahel keerukalt põimunud. Seetõttu on ülemises Metazoas primaarsed kihid taandatud idukihtide tasemele; neid ei ole enam täiskasvanul, vaid need säilivad embrüo esmaste kihtidena, tekitades täiskasvanud organismi teatud rakusüsteeme, kudesid ja elementaarorganeid. Need idukihid jäävad aga kõigis Metazoades üksteisega homoloogseks kõikjal, välja arvatud täiskasvanud käsnades, säilitades samad vastastikuse asukoha ja tulevase tähtsuse iseloomulike tunnuste põhikomplektid.
Järeldus
Niisiis, idukihid ei ole väljamõeldud mõiste, nad on tõesti olemas, nad näitavad teatud tüüpi rakumaterjali esmast diferentseerumist metazoa munast arenemise ajal. Idukihtide püsivust enamiku loomade arengus saab seletada ainult "ajalooliste traditsioonide", st kokkuvõtete olemasoluga. Kuid idukihte ei tohiks pidada millekski stabiilseks ja muutumatuks; ei tohiks unustada mis tahes ontogeneetiliste protsesside võimalikke evolutsioonilisi transformatsioone, sealhulgas idukihtide arengut.
Kirjandus
1. Ivanova-Kazas O. M., Krichinskaya E. B. Kursus selgrootute loomade võrdlevast embrüoloogiast. L. Kirjastus Leningrad. Ülikool, 1988.
2. http:///biologia/26-zarodyshevye-listki. html
3. Suur Nõukogude Entsüklopeedia, TSB
Mis on idukihid või -kihid? Mis on selle termini tähendus? Artiklis antakse lühike teave nende eraldatud rakurühmade kohta, mis esinevad kõigis loomastiku esindajate embrüodes teatud embrüonaalse arengu staadiumis.
Ajaloost
Veel 18. sajandi 60ndatel jälgis ja hiljem kirjeldas Saksa ja Vene füsioloog Caspar Friedrich Wolf ühe idukihi moodustumist ja muutumist sooletoruks. Esmakordselt avastas ja kirjeldas kõik kolm idukihti Peterburi Keiserliku Teaduste Akadeemia akadeemik (1821), loodusteadlane, embrüoloog ja paleontoloog Christian Heinrich Pander. Ta uuris nende struktuuri, uurides ka kana embrüot. Lisaks avastas sama akadeemia akadeemik Karl Baer idukihtide olemasolu ka teiste loomade – kalade, roomajate, kahepaiksete – embrüodes. Tänu nende teadlaste töödele anti nende struktuuride uurimisele tõuge.
Idukihtide moodustumine
Sügoot (looma viljastatud munarakk) hakkab jagunema. Embrüonaalse arengu varases staadiumis jagunevad rakud intensiivselt mitoosi teel, moodustades sfäärilise struktuuri - morula ja seejärel - blastula. Selle erinevus morulast seisneb selles, et selles etapis lahknevad rakud (neid nimetatakse blastomeerideks) keskelt perifeeriasse ja keskele moodustub nn blastodermi vesiikul. Seetõttu on Blastula ühekihiline embrüo.

Pärast selle loomamaailma esindajate embrüonaalse arengu perioodi, mida nimetatakse purustamiseks, lõppu algab gastrulatsioonifaasi pööre. Erinevus nende ontogeneesi etappide vahel on kardinaalne. Esimesel juhul jaguneb viljastatud munarakk paljudeks blastomeerideks (väiksemateks rakkudeks), samas kui mass ja maht ei muutu. Purustamise peamine tähendus on embrüo üleminek ühest rakust paljurakulisusele. Gastrulatsioon, mis toimub pärast purustamist, tähendab rakkude diferentseerumist. Selles etapis ilmuvad nn idukihid. Need on teatud rakurühmad, millest hiljem moodustuvad teatud kuded ja elundid.
Erinevused idukihtides
Embrüo struktuur gastrulatsiooni staadiumis ja sellele eelnev on näidatud alloleval pildil. Gastrulatsioonile järgnevas staadiumis, mida nimetatakse neurulaks, moodustub närviplaat, akordi alge, epiteel ja sooled. Keha tagumine ja eesmine osa muutuvad eristatavaks.

Gastrulatsiooni ajal, nagu eespool mainitud, ei toimu mitte ainult rakkude paljunemine, vaid ka nende kasv ja suunatud liikumine, mis viib hiljem selgelt väljendunud diferentseerumiseni. Seotud rakkude rühmad ühendatakse eraldi rakkude kihtideks, välisteks ja sisemisteks. Neid nimetatakse ektodermiks ja endodermiks.
Käsnad ja koelenteraadid (meduusid, korallid, ktenofoorid) arendavad ainult neid kahte idukihti. Kõrgematel loomadel moodustub neist kolm: mainitud ektoderm ja endoderm, samuti keskmine leht - mesoderm.
Nende erinevus seisneb eelkõige funktsioonides, aga ka selles, milliseid elundeid ja kudesid nad tekitavad. Neid käsitletakse üksikasjalikumalt allpool.

ektoderm
Sugurakkude välimine kiht vastutab motoorsete, sensoorsete ja sisemiste funktsioonide eest. Sellest arenevad hiljem närvisüsteemi organid. Lisaks areneb ektodermist nahk ja kõik, mis loomadel peal on: kaitsesoomused, küünised, küüned, suled, kilbid jne, aga ka hambaemail.
See selgroogsete idukiht koosneb kolmest osast: välimine, samuti neuraaltoru ja närvihari. Kaks viimast komponenti on tuntud ka kui neuroektodermid. Kanada embrüoloogi Brian Halli ettepanekul on närviharja alates 2000. aastast paljudes väljaannetes nimetatud neljandaks idukihiks.
Endoderm
Idukiht, millest osaliselt moodustuvad siseorganid. See on seedesüsteem, sealhulgas näärmed (kõhunääre, maks). Endodermist (kaladel, lõpustel ja ujupõies) arenevad ka hingamiselundid.
mesoderm
Sugurakkude keskmine kiht, mis on iseloomulik ainult kõrgematele loomadele. Vastutab troofiliste ja tugifunktsioonide täitmise eest. See arendab luid ja lihaseid, kõhre, nookordi, eritusorganeid, samuti reproduktiiv- ja vereringeelundeid.
Lõpuks
Artiklis kirjeldati lühidalt loomade idukihte, nende funktsioone, loetleti organid ja süsteemid, mis arenevad mesodermist, ektodermist, endodermist.
Huvitav fakt on see, et kõigis loomamaailma esindajates on enamikus elundites kudesid 2–3 neist struktuuridest.