Mikroorganizmaların ana formlarına göre sınıflandırılması. Bakteriler, çeşitlilikleri. Yapı. canlılık. Temel terimler: bakteri
Bakterilerin beslenmesi.
Gıda.
pasif difüzyon
Kolaylaştırılmış difüzyon
aktif taşımacılık
İlk durumda, besin molekülü, belirli bir sitoplazmik zar geçirgenliği ile etkileşime giren bir periplazmik boşluk proteini ile bir kompleks oluşturur. Sitoplazmik membrandan enerjiye bağlı penetrasyondan sonra, "substrat - periplazmik protein - permeaz" kompleksi, substrat molekülünün salınımı ile ayrışır.
Taşınan maddenin kimyasal modifikasyonu ile aktif taşıma sırasında, olaylar zinciri şunları içerir: (1) membran enzim-2'nin sitoplazmadan fosfoenolpiruvat ile fosforilasyonu; (2) fosforile enzim-2 substrat molekülleri tarafından sitoplazmik membranın yüzeyine bağlanma; (3) substrat molekülünün sitoplazmaya enerjiye bağlı taşınması; (4) bir fosfat grubunun bir substrat molekülüne transferi; (5) sitoplazmada "substrat-enzim" kompleksinin ayrılması. Fosforilasyon nedeniyle, substrat molekülleri hücrelerin sitoplazmasında birikir ve onları terk edemezler.
Bakterilerin gıda türüne göre sınıflandırılması.
Besin alımı yoluyla bakteriler sınıflandırılır holofitler ve Holozoik. Holofit bakteri (Yunancadan. hololar- tamamlayın ve bitkisel- bitkilerle ilgili), substratları parçalayan enzimleri çevreye salamazlar, bunun sonucunda besinleri yalnızca çözünmüş, moleküler bir biçimde tüketirler. Holozoik bakteri (Yunancadan. hololar- tamamlayın ve zoikolar- hayvanlarla ilgili), aksine, dış beslenme sağlayan bir ekzoenzim kompleksine sahiptirler - substratların bakteri hücresi dışındaki moleküllere parçalanması. Bundan sonra, besin molekülleri Holozoik bakterilerin içine girer.
Karbon kaynağına göre bakterilerden izole edilmiş ototroflar ve heterotroflar. ototroflar (Yunancadan. arabalar- kendim, ganimet- gıda) karbondioksit (CO 2), karbon içeren tüm maddelerin sentezlendiği bir karbon kaynağı olarak kullanılır. İçin heterotroflar (Yunanca geteros'tan - başka ve ganimet- gıda) karbon kaynakları moleküler formda (karbonhidratlar, polihidrik alkoller, amino asitler, yağ asitleri) çeşitli organik maddelerdir. En yüksek heterotrofi derecesi, yalnızca diğer canlı hücrelerin (örneğin, riketsiya ve klamidya) içinde yaşayabilen prokaryotlarda bulunur.
Azot kaynağına göre prokaryotlar 3 gruba ayrılır: 1) nitrojen sabitleyen bakteriler (moleküler nitrojeni atmosferik havadan özümler); 2) amonyum tuzlarından, nitritlerden veya nitratlardan inorganik nitrojen tüketen bakteriler; 3) organik bileşiklerde bulunan azotu özümseyen bakteriler (amino asitler, pürinler, pirimidinler, vb.).
Enerji kaynağına göre bakteriler ikiye ayrılır fototroflar ve kemotroflar. fototrofik bakteri bitkiler gibi güneş enerjisini kullanabilirler. Fototrofik prokaryotlar insanlarda hastalığa neden olmaz. kemotrofik bakteri redoks tepkimelerinde enerji alır.
Elektron donörlerinin doğası gereği litotroflar(Yunancadan. litolar- taş) ve organotroflar. saat litotroflar (kemolitotroflar ) inorganik maddeler elektron verici görevi görür (H 2, H 2 S, NH 3, kükürt, CO, Fe 2+, vb.). elektron vericileri organotroflar (kemoorganotroflar ) organik bileşiklerdir - karbonhidratlar, amino asitler vb.
İnsanlar için patojenik bakterilerin çoğu, kemoorganotrofik (kemoheterotrofik) bir beslenme tipine sahiptir; kemolitotrofik (kemoototrofik) tip daha az yaygındır.
Organik bileşikleri sentezleme yeteneği ile kemotrofik bakteriler şu şekilde sınıflandırılır: prototroflar, oksotroflar ve hipotroflar. prototrofik bakteri gerekli tüm organik maddeleri glikoz ve amonyum tuzlarından sentezler. bakteri denir oksotroflar belirtilen bileşiklerden herhangi bir organik madde sentezleyemezlerse. Aşırı derecede metabolik aktivite kaybı, yetersiz beslenme olarak adlandırılır. hipotrofik bakteri konakçının yapılarını veya metabolitlerini yeniden düzenleyerek hayati aktivitelerini sağlarlar.
Bakterilerin tam teşekküllü yaşamı için karbon ve azotun yanı sıra kükürt, fosfor ve metal iyonları gereklidir. Kükürt kaynakları amino asitler (sistein, metionin), vitaminler, kofaktörler (biyotin, lipoik asit vb.), sülfatlardır. Fosfor kaynakları nükleik asitler, fosfolipitler, fosfatlardır. Yeterince yüksek konsantrasyonlarda bakterilerin magnezyum, potasyum, kalsiyum, demire ihtiyacı vardır; çok daha küçük miktarlarda - çinko, manganez, sodyum, molibden, bakır, nikel, kobalt.
büyüme faktörleri- Bakterilerin kendi kendilerine sentezleyemedikleri ancak çok ihtiyaç duydukları maddelerdir. Amino asitler, azotlu bazlar, vitaminler, yağ asitleri, demir porfirinler ve diğer bileşikler büyüme faktörleri olarak işlev görebilir. Bakterilerin hayati aktivitesi için en uygun koşulları yaratmak için, besin ortamına büyüme faktörleri eklenmelidir.
Metabolizma, enerji dönüşümü
A) Yapıcı metabolizma.
Bakteriyel beslenmenin zorunlu bir aşaması, besinlerin asimilasyonu, yani hücresel bileşenlerin ve yapıların çoğaltılması için sentetik reaksiyonlara değiştirilmiş veya değiştirilmiş bir biçimde dahil edilmeleridir.
Protein metabolizması bakterilerde 3 aşamada ilerleyebilir: birincil protein yıkımı, ikincil protein yıkımı ve protein sentezi. Protein moleküllerinin peptonlara birincil parçalanması, bakteriler tarafından çevreye salınan ekzoenzimler - ekzoproteazlar tarafından gerçekleştirilir. İkincil bozunma, tüm bakterilerin sahip olduğu endoenzimlerin (endoproteazlar) etkisi altında meydana gelir. Bu süreç bakteri hücresinin içinde gerçekleşir ve peptitlerin kendilerini oluşturan amino asitlere parçalanmasından oluşur. İkincisi değişmeden kullanılabilir veya amonyak, indol, hidrojen sülfür, keto asitler, alkol, karbon dioksit ve diğerlerinin ortaya çıkmasına neden olan kimyasal dönüşümlere (deaminasyon, dekarboksilasyon, vb.) tabi tutulabilir. bakteriyolojide tanısal öneme sahiptir.
Protein parçalanma reaksiyonları ile birlikte sentez reaksiyonları meydana gelir. Bazı bakteriler dışarıdan beslenme sonucu elde edilen hazır amino asitlerden proteinler oluştururken, diğer bakteriler azot ve karbon içeren basit bileşiklerden bağımsız olarak amino asitleri sentezler. Amino asitlerin sentezi, aminasyon, transaminasyon, amidasyon, karboksilasyon reaksiyonlarında gerçekleştirilebilir. Çoğu prokaryot, hücresel proteinleri oluşturan tüm amino asitleri sentezleyebilir. Amino asitlerin biyosentezinin bir özelliği, ortak biyosentetik yolların kullanılmasıdır: trikarboksilik asit döngüsü, glikoliz, oksidatif pentoz fosfat yolu, vb. Amino asitlerin sentezi için ana başlangıç bileşiği piruvat ve fumarattır.
Karbonhidrat metabolizması ototroflarda ve heterotroflarda farklılık gösterir (Şema 1). Ototrofik bakteriler, gerekli tüm karbonhidratları karbondioksitten sentezler. Heterotrofik bakterilerde karbonhidrat oluşumu için hammaddeler şunlar olabilir: (1) bir, iki ve üç karbonlu bileşikler; ve (2) polisakaritler (nişasta, glikojen, selüloz). İkincisini bölmek için, birçok heterotrofik bakteri, glikoz, maltoz, fruktoz vb. oluşturmak üzere polisakkaritleri hidrolize eden ekzoenzimlere (amilaz, pektinaz, vb.) sahiptir.
Ototrofik bakterilerde, Calvin döngüsünde, ters yönde giden glikoliz reaksiyonlarına dahil olan karbondioksitten ribuloz fosfat-fosfor-gliserik asit oluşur. Ters sentezin son ürünü glikozdur.
Heterotrofik bakteriler bir, iki ve üç karbonlu bileşiklerden glikoz oluşturur ve bunları ters glikoliz reaksiyonunda da içerir. Bazı glikoliz reaksiyonlarının geri döndürülemez olması nedeniyle, heterotroflar, katabolik yolun geri dönüşü olmayan reaksiyonlarını atlamalarına izin veren özel enzimatik reaksiyonlar oluşturmuştur.
Polisakkaritler heterotrofik bakteriler tarafından parçalandığında, ortaya çıkan disakkaritler hücrelere girer ve maltoz, sakaroz ve laktozun etkisi altında hidrolize uğrar ve monosakkaritlere parçalanır, bunlar daha sonra fermente edilir veya şeker dönüşüm reaksiyonlarına dahil edilir.
Lipid metabolizması. Hem eksojen lipidler hem de interstisyel metabolizmanın amfibolitleri, bakterilerde lipidlerin oluşumu için başlangıç materyalleri olarak hizmet edebilir. Eksojen lipidler, bakteriyel lipazlara ve diğer lipolitik enzimlere maruz kalır. Birçok bakteri türü, plastik malzeme ve enerji kaynağı olarak hizmet eden gliserolü emer. Lipid sentezi için endojen kaynaklar, asetilkoenzim A, propionil-APB, malonil-APB (ACP - asetil-taşıyıcı protein), fosfodioksiaseton vb. olabilir.
Çift sayıda karbon atomlu yağ asitlerinin sentezi için ilk substrat, tek sayıda karbon atomlu yağ asitleri için asetilkoenzim A, propionil-APB ve malonil-APB'dir. Aerobik prokaryotlarda asit molekülünde çift bağların oluşumu moleküler oksijen ve enzim desatürazın katılımıyla gerçekleşir. Anaerobik prokaryotlarda, bir dehidrasyon reaksiyonunun bir sonucu olarak sentezin başlarında çift bağlar eklenir. Fosfolipidlerin sentezi için ilk substrat, indirgenmesi 3-fosforogliserol oluşumuna yol açan fosfodioksiasetondur (glikolitik yolun bir ara bileşiği). Daha sonra ikincisine APB ile bir kompleks şeklinde 2 yağ asidi kalıntısı eklenir. Reaksiyon ürünü, aktivasyonu CTP ile serin, inositol, gliserol veya başka bir bileşiğin fosfat grubuna bağlanması, karşılık gelen fosfolipidlerin sentezine yol açan fosfatidik asittir.
Yağ asitleri için oksotropik ve hipotrofik olan mikroorganizmalar (örneğin, mikoplazmalar) onları hazır olarak konakçı hücrelerden veya bir besin ortamından alırlar.
mononükleotid değişimi. Purin ve pirimidin mononükleotitleri, DNA ve RNA'nın temel bileşenleridir. Birçok prokaryot, besin ortamında bulunan hazır pürin ve pirimidin bazlarını, bunların nükleozidlerini ve nükleotidlerini hem kullanabilmekte hem de düşük moleküler ağırlıklı maddelerden sentezleyebilmektedir. Bakteriler, ekzojen pürin ve pirimidin türevlerinin karşılıklı dönüşümlerinin aşağıdaki aşamalarını katalize eden enzimlere sahiptir: azotlu baz - nükleosit - nükleotid (mono-di-trifosfat).
Purin ve pirimidin mononükleotitlerinin sentezi yeni bağımsız yollarla gerçekleştirilir. Pürin nükleotitlerinin sentezi sırasında, ardışık enzimatik reaksiyonların bir sonucu olarak, pürin halkasının kimyasal modifikasyonları ile adenil (AMP) ve guanilik (GMP) asitlerin sentezlendiği inosinik asit oluşur. Pirimidin nükleotitlerinin sentezi, dekarboksilasyonu uridilik asit (UMP) veren orotidilik asit oluşumu ile başlar. İkincisinden, asilasyonu CTP oluşumuna yol açan UTP oluşur.
Deoksiribonükleotitler, karşılık gelen ribonükleotitlerin difosfat veya trifosfat seviyesinde indirgenmesinin bir sonucu olarak oluşur. DNA'ya özgü bir nükleotidin, timidilik asidin sentezi, deoksiuridilik asidin enzimatik metilasyonu ile gerçekleşir.
İyon değişimi. Mineral bileşikler - iyonlar, NH 3 + , K + , Mg 2+ , Fe 2+ , SO 4 2- , PO 4 3- ve diğer bakteriler hem serbest halde hem de diğer organik maddelerle ilişkili halde ortamdan elde edilir. maddeler. Katyonlar ve anyonlar, § 3'te açıklanan çeşitli şekillerde bakteri hücresine taşınır. İyonların bakteri hücresine girme hızı, ortamın pH'ından ve mikroorganizmaların kendilerinin fizyolojik aktivitelerinden etkilenir.
B) Bakterilerin solunumu (enerji metabolizması).
Tüm yaşam süreçleri enerjiye bağlıdır, bu nedenle enerji elde etmek prokaryotların metabolizmasının son derece önemli bir yönüdür. Anaerobik ve aerobik solunumdan enerji alırlar.
Nefes, veya biyolojik oksidasyon, makroerjik bileşiklerde enerji birikiminin eşlik ettiği bir verici maddeden bir alıcı maddeye elektron transferinin katabolik bir sürecidir. . Solunum, katabolik reaksiyonlar sürecinde gerçekleştirilir, bunun sonucunda karmaşık organik maddeler, parçalanır, enerji verir ve basit bileşiklere dönüşür. Makroerjik maddelerde (ATP, GTP, UTP vb.) biriken enerji anabolik reaksiyonlarda kullanılır.
Mikroorganizmalar solunum şekline göre şu şekilde sınıflandırılır: zorunlu (katı) aeroblar, zorunlu anaeroblar ve fakültatif anaeroblar.
zorunlu aeroblar serbest oksijene ihtiyaç duyar. Organik bileşikler (karbonhidratlar, yağlar, proteinler) kemoorganotrofik aeroblarda insanlar için patojenik elektron vericilerdir ve moleküler oksijen bir elektron alıcısıdır. Enerjinin kemoorganotrofik aeroblarda ATP formunda depolanması, elektron donörlerinin oksidatif fosforilasyonu sırasında meydana gelir. Aeroblar, solunum sırasında oluşan toksik oksijen radikallerini etkisiz hale getiren enzimlerin (katalaz, süperoksit dismutaz, peroksidaz) yanı sıra sitokromlara (elektron transferine katılır) sahiptir. Süperoksit dismutaz, en toksik metaboliti, H 2 O 2'deki süperoksit radikalini etkisiz hale getirir. Katalaz enzimi H 2 O 2 yi H 2 O ve O 2 ye dönüştürür.
Özel bir aerob grubu mikroaerofilik bakteri enerji için oksijene ihtiyaç duymalarına rağmen, artan CO2 içeriği ile daha iyi büyürler, örneğin, cinsin bakterileri kampilobakter ve Helikobakter.
zorunlu anaeroblar serbest oksijene ihtiyaç duymazlar, tam tersine, oksijenin küçük miktarlarda bile onlar üzerinde toksik etkisi vardır. İnsanlar için patojen olan anaerobik kemoorganotroflardaki elektron donörleri, çeşitli organik bileşiklerdir (esas olarak karbonhidratlar). Kemoorganotrofik anaeroblardaki elektron alıcısı organik oksijen içeren bileşiklerdir - asitler veya ketonlar, yani elektron alıcısı organik parça ile ilişkili oksijendir. Bu prokaryotlarda enerjinin depolanması, substrat fosforilasyonu sırasında gerçekleşir. Zorunlu anaeroblar, kural olarak, oksijen radikallerini (katalaz ve süperoksit dismutaz-negatif) etkisiz hale getiren sitokromlara ve enzimlere sahip değildir.
İnsanlar için patojenik olmayan kemolitotrofik anaeroblarda elektron alıcısı inorganik oksijen içeren bileşiklerdir - nitratlar, sülfatlar, karbonatlar.
Özel bir anaerob grubu, atmosferik oksijen varlığında büyüyebilen, ancak elektron alıcısı olarak kullanmayan (örneğin, laktik asit bakterileri) aerotolerant bakterilerdir. Aerotolerant katalaz ve süperoksit dismutaz prokaryotları pozitiftir.
Fakültatif anaeroblar hem oksijenli hem de anoksik ortamlarda bulunabilirler. Elektron vericileri organik maddelerdir; elektron alıcıları, çevresel koşullara bağlı olarak, organik ve inorganik bileşiklerde moleküler veya oksijene bağlıdır. Fakültatif anaeroblar hem oksidatif hem de substrat fosforilasyonu sırasında enerji biriktirebilir. Aeroblar gibi, bu bakteri grubu da sitokromlara ve antioksidan savunma enzimlerine sahiptir.
Enerji elde etmek için ana substrat, farklı solunum tiplerinin kemoheterotrofik prokaryotlarında asetilkoenzim A'ya (“aktive edilmiş asetik asit”) katabolize edilebilen karbonhidratlardır. Asetil koenzim A aynı zamanda metabolizmalarının ara ürünlerinden biri olduğu için lipitler ve proteinler enerji substratları olarak hareket edebilir (Şema 2).
Kemoorganotrofik prokaryotlarda karbonhidrat katabolizması şunları içerir: (a) anaerobik süreçler—glikoliz, pentoz fosfat yolu ve ketodeoksifosfoglukonat yolu; (b) aerobik süreç - trikarboksilik asit döngüsü (Krebs döngüsü). Anaerobik süreçler tüm prokaryotlarda gerçekleşirken, aerobik süreç sadece zorunlu aerobların ve fakültatif anaerobların karakteristiğidir. Anaerobik yollar, substrat fosforilasyonuna dayanırken, aerobik süreçler oksidatif fosforilasyona dayanır.
Kavramların tanımı.
Sterilizasyon, dezenfeksiyon ve antiseptikler, modern tıp ve özellikle cerrahi pratiğin ayrılmaz parçalarıdır. Bu yöntemlerin prensiplerini ve pratik uygulamalarını anlamak esastır çünkü birçok potansiyel olarak patojenik mikroorganizma, konakçı dışında uzun süre hayatta kalabilir, fiziksel ve kimyasal dezenfektanlara karşı yüksek direnç gösterebilir ve kişiden kişiye nispeten kolay yayılabilir.
antiseptikler- kimyasal yöntemlerle patojenik veya fırsatçı mikroorganizmaların büyümesinin yok edilmesi veya önlenmesi. Bu terim genellikle bir kimyasal preparasyonun canlı dokulara harici olarak uygulanmasına atıfta bulunmak için kullanılır.
antiseptik- bir mikroorganizmanın büyümesini engelleyen veya yok eden bir madde (bakteri sporları üzerinde etki olmaksızın). Terim, canlı dokular üzerinde topikal etki için kullanılan maddelere özeldir.
Asepsi sepsisin yokluğu anlamına gelir, ancak genel olarak bu terim herhangi bir canlı organizmanın yokluğunu vurgulamak için kullanılır. aseptik yöntemler canlı organizmaları ortadan kaldırmak ve bunlar tarafından yeniden kontaminasyonu önlemek için tasarlanmış herhangi bir prosedür anlamına gelir. Modern cerrahi ve mikrobiyolojik teknikler aseptik prosedürlere dayanmaktadır.
biyosit- sporlar da dahil olmak üzere hem patojenik hem de patojenik olmayan tüm canlı mikroorganizmaları öldüren bir madde.
biyostat- Mikroorganizmaların büyümesini engelleyen, ancak onları mutlaka öldürmeyen bir ajan.
dekontaminasyon- kantitatif belirleme olmaksızın mikroorganizmaların uzaklaştırılması. Bu terim görecelidir; mikropların nihai olarak uzaklaştırılması sterilizasyon veya dezenfeksiyon ile gerçekleştirilebilir.
dezenfeksiyon- sporlar hariç tüm patojenleri azaltan veya ortadan kaldıran bir süreç.
Antiseptik- mikroorganizmaları, özellikle patojenleri yok eden bir madde. Mikrop öldürücü sporları yok etmez.
sanitasyon- mikrobiyal kontaminasyonun “güvenli” bir düzeye indirildiği bir yöntem. Bu yöntem daha önce cansız nesneleri "arındırmak" için kullanılıyordu.
Sterilizasyon- her türlü mikrobiyal yaşamın tamamen yok edilmesi veya yok edilmesi için fiziksel faktörlerin ve (veya) kimyasalların kullanılması.
Sterilizasyon.
Sterilizasyon, tüm mikroorganizmaların ve sporlarının yok edilmesi veya uzaklaştırılması (filtreleme yoluyla) olarak tanımlanır. Sterilizasyon genellikle ısı kullanılarak gerçekleştirilir. Bir mikrobiyoloji laboratuvarının günlük rutin çalışmalarından biri olan sterilizasyon, kültürlerin, ekipmanların, gereçlerin ve ortamın sadece gerekli mikroorganizmaların büyümesini desteklerken diğer mikropları yok etmesini sağlamak için gerekli bir yöntemdir. Bu tür sterilizasyon türleri vardır: brülör alevinde kalsinasyon, kaynatma, akan buharla hareket, otoklavda basınç altında buhar, kuru ısı, pastörizasyon, tyndalizasyon, kimyasal, soğuk (mekanik) sterilizasyon.
Sterilizasyon yöntemlerinin seçimi.
Sterilizasyon yöntemlerini seçerken aşağıdaki gereksinimler dikkate alınmalıdır:
1. Aktivite: bakterisit, spor öldürücü, tüberkülosidal, mantar öldürücü ve virüs öldürücü.
2. İşlem hızı: sterilizasyon mümkün olduğunca çabuk yapılmalıdır.
3. Geçirgenlik: Sterilize edici ajanlar, ambalajın içinden ve aletin iç kısmına nüfuz edebilmelidir.
4. Uyumluluk: Birkaç kez sterilize edilen malzemelerin yapısında veya işlevinde herhangi bir değişiklik olmamalıdır.
5. Toksik olmayan: İnsan sağlığı ve çevre için herhangi bir tehdit olmamalıdır.
6. Organik materyalin kalıcılığı: Organik materyalin varlığında sterilizasyon etkinliği azalmamalıdır.
7. Uyarlanabilirlik: Büyük ve küçük hacimli sterilize edilmiş malzeme için kullanım yeteneği.
8. Zaman içinde kontrol: işleme döngüsü kolay ve doğru bir şekilde kontrol edilmelidir.
9. Fiyat: makul ekipman, kurulum ve işletme maliyeti.
 Fiziksel sterilizatörler
Fiziksel sterilizatörler
ıslak ılık, buharlı otoklavlama işlemi sırasında oluşan, klinik mikrobiyoloji laboratuvarlarında kullanılan ana sterilizasyon ajanıdır. Otoklavlar, kültür ortamını, refrakter malzemeleri sterilize etmek ve bulaşıcı atıkları tedavi etmek için kullanılır. Bir buhar sterilizatörü veya otoklav, yüksek sıcaklıklar oluşturmak için doymuş buhar kullanan yalıtılmış basınçlı bir odadır (Şekil 1). Hava, kütle yer değiştirmesi veya vakumla hazneden çıkarılır. Ağırlıkça ikameli en yaygın kullanılan otoklavlar. Daha ağır havayı değiştirmek için hazneye daha hafif buhar verilir. Basınçlı buhara kısa süre maruz kalmak bakteri sporlarını yok edebilir. Kültür ortamının ve diğer malzemelerin rutin sterilizasyonu için, maruz kalma süresi 121°C'de 15 dakika ve 1 santimetre kare başına 1.5 kg'lık bir basınçtır. Enfeksiyöz atıklar için maruz kalma süresi 30-60 dakikaya çıkarılır. Doğru zaman ve sıcaklığın yanı sıra buharla doğrudan temas sterilizasyon için çok önemlidir. Enfeksiyöz malzeme ile uğraşırken, buharın atığa maksimum penetrasyonu sağlanmalıdır. Bu tür malzeme 132ºС sıcaklıkta işlenmelidir. Ayrışamayan antineoplastikler, toksik kimyasallar ve radyoizotoplar ve stabil olmayan kimyasallar, ısıya maruz kaldıklarında buharlaşıp oda içerisinde yayılabileceğinden otoklavlanmamalıdır.
 Kuru ısı sterilizasyonu Hasar olasılığı veya malzemenin buhar geçirimsizliği nedeniyle buharla sterilize edilemeyen malzemeler için kullanılır. Kuru ısı, nemli ısıdan daha az etkilidir ve daha uzun maruz kalma süreleri ve daha yüksek sıcaklıklar gerektirir. Kuru ısı ile sterilizasyon genellikle kuru ısı kabininde gerçekleştirilir (Şekil 2). Kuru ısı sterilizasyonunun mekanizması oksidatif bir süreçtir. Kuru ısı sterilizasyonunun kullanıldığı malzeme örnekleri, yağlar, tozlar, keskin aletler ve cam eşyalardır. Enfeksiyöz atıkların arıtılmasında alternatif yöntemler olarak kuru ısı veya termal inaktivasyon-sterilizasyon kullanılmaktadır.
Kuru ısı sterilizasyonu Hasar olasılığı veya malzemenin buhar geçirimsizliği nedeniyle buharla sterilize edilemeyen malzemeler için kullanılır. Kuru ısı, nemli ısıdan daha az etkilidir ve daha uzun maruz kalma süreleri ve daha yüksek sıcaklıklar gerektirir. Kuru ısı ile sterilizasyon genellikle kuru ısı kabininde gerçekleştirilir (Şekil 2). Kuru ısı sterilizasyonunun mekanizması oksidatif bir süreçtir. Kuru ısı sterilizasyonunun kullanıldığı malzeme örnekleri, yağlar, tozlar, keskin aletler ve cam eşyalardır. Enfeksiyöz atıkların arıtılmasında alternatif yöntemler olarak kuru ısı veya termal inaktivasyon-sterilizasyon kullanılmaktadır.
Pastörizasyon, maddeyi 15 saniye boyunca hızla 71.1°C'ye ısıtarak ve ardından hızlı bir şekilde soğutarak patojenleri yok eder. Pastörizasyon sterilizasyon değildir çünkü tüm mikroorganizmalar buna duyarlı değildir. Bu yöntem, gastrointestinal tüberküloz ve Q ateşi gibi hastalıkların gıda kaynaklı bulaşmasını ortadan kaldırdı.
Tyndalizasyon solüsyonlardaki tüm bakterileri öldürmek için kullanılabilen aralıklı bir ısı sterilizasyon yöntemidir. Büyüyen bakteriler kısa kaynamalarla (5 dakika boyunca 1 saat içinde 5 kez) kolayca öldürüldüğünden, yapılması gereken tek şey, ısı sporların olgunlaşmasını bozana kadar çözeltinin belirli bir süre beklemesine izin vermektir. ısıya karşı dirençlerinde önemli bir kayıp.
filtreleme solüsyonlardan, havadan ve diğer gazlardan mikropları ve mikroskobik partikülleri uzaklaştırmak için kullanılan bir işlemdir. Laboratuvarda filtrasyon ile sterilizasyonun en yaygın kullanımı teşhis, kültür ortamı, doku kültürü ortamı, serum, serum bileşenleri içeren solüsyonların işlenmesidir. Filtrasyonun diğer bir yaygın uygulaması, hava ve gazların sterilizasyonudur. Gözenek çapı (yaklaşık 12 ila 0,22 µm) ile ayırt edilen ve mekanik ayırma için kullanılan plastik veya kağıt membran filtreler, aynı zamanda, ıslatılmış bir yüzeye yerleştirildiğinde doğrudan filtre üzerinde mikroskobik inceleme veya yetiştirme için sıvılardan mikrop toplamaya da hizmet eder. orta.
ultraviyole ışınımı hücresel nükleik asit üzerinde etkili olan bir tür elektromanyetik dalga radyasyonudur. Mikroorganizmalar, 254 nm dalga boyuna sahip ultraviyole ışınlarının etkisine karşı oldukça hassastır. Ultraviyole ışık en yaygın olarak havadaki veya yüzeylerdeki mikroorganizmaları öldürmek için kullanılır. Diğer kullanımlar, farmasötik uygulamalar için belirli kimyasalların ve plastiklerin soğuk sterilizasyonu, hücre kültürleri için serum sterilizasyonu ve su dezenfeksiyonudur. Bir sterilizatör olarak ultraviyole ışınımının önemli bir dezavantajı, malzemelere nüfuz edememesidir.
iyonlaştırıcı radyasyon elektromanyetik spektrumda, mikroorganizmalar üzerinde öldürücü bir etkiye sahiptir. Bu spektrum mikrodalgaları, γ-ışınlarını, X-ışınlarını ve elektron akışını içerir. İyonlaştırıcı radyasyonun öldürücü etkisi, hedef molekül üzerindeki doğrudan etki nedeniyle meydana gelir ve bunun sonucunda moleküle enerji aktarılır; ve dolaylı etki nedeniyle - radikallerin difüzyonu.
ultrasonik enerji düşük frekanslı sulu çözeltilerdeki mikroorganizmaları inaktive eder. Sonikasyonun fiziksel etkisi kavitasyondan kaynaklanır. Ultrasonik temizleyiciler ve diğer cihazlar genellikle aletleri temizlemek için kullanılır ancak sterilizatör olarak kabul edilmez. Bununla birlikte, ultrasonun kimyasal işlemle kombinasyonu mikroorganizmaları öldürür.
Kimyasal sterilizatörler
2 % glutaraldehit Sıvı bir kimyasal sterilizatör olarak, daha önce ısıtma veya ışınlama ile sterilize edilemeyen tıbbi ve cerrahi malzemeleri işlemek için yaygın olarak kullanılıyordu. Glutaraldehit ayrıca aşıların hazırlanmasında da kullanılır.
Dezenfeksiyon.
Dezenfeksiyon kimyasal yöntemlerle veya kaynatılarak yapılabilir. Kaynatma, otoklav yoksa iğne ve şırınga gibi aletleri dezenfekte etmek için etkili bir yöntemdir. Önceden temizlenmiş tıbbi aletler 20 dakika kaynatılmalıdır. Yüksek sıcaklıklardan zarar görebilecek ısıya duyarlı ekipmanlar için kimyasal dezenfeksiyon kullanılmaktadır. Klor bileşenleri, etil ve izopropil alkol, kuaterner amonyum bileşenleri ve glutaraldehit gibi kimyasal dezenfektanlar yaygın olarak kullanılmaktadır.
Kimyasal dezenfektanlar.
Alkol (etil ve izopropil) Suda %60-85 konsantrasyona kadar çözülür, dezenfeksiyonda çok etkilidir. Alkoller bakterisit, mantar öldürücü ve tüberküloziddir, ancak sporları etkilemez. Etil alkol, izopropil alkolden daha geniş bir virüs öldürücü aktivite spektrumuna sahiptir, bu nedenle lipofilik ve hidrofilik virüslere karşı daha etkilidir.
Çözüm %37 formaldehit formalin adı verilen , sterilizatör olarak kullanılabilirken, %3-8 konsantrasyonları dezenfektan olarak kullanılabilir.
Fenol Toksisitesi, tümör gelişimini indükleme ve korozyona neden olma özelliğinden dolayı saf haliyle dezenfektan olarak kullanılmaz. Aromatik halkadaki hidrojen atomlarından birinin yerini fonksiyonel bir grubun (klor, brom, alkil, benzil, fenil, amil) aldığı fenol türevleri dezenfektan olarak yaygın olarak kullanılmaktadır. Bu ikame, fenolün dezavantajlarını azaltır. Fenolün bileşenleri, enzim sistemlerinin inaktivasyonu, proteinlerin çökelmesi ve hücre duvarı ve zarının bozulması nedeniyle mikropları öldürür. Genellikle %2-5'lik konsantrasyonlar kullanılır, daha düşük konsantrasyonlar daha uzun süre maruz kalmayı gerektirir.
Halojenler. Laboratuvar uygulamalarında dezenfeksiyon için sadece klor ve iyot kullanılmaktadır. Klorun güçlü bir oksitleyici ajan olması nedeniyle mikropları oksidasyon yoluyla öldürdüğüne inanılır. İyotun, amino asitlerin N-H ve S-H grupları ile ve ayrıca amino asit tirozinin fenolik grubu ve doymamış yağ asitlerinin karbon-karbon çift bağları ile reaksiyona girerek mikroorganizmaları öldürdüğüne inanılmaktadır. Geleneksel tedavi, 60-80ºC sıcaklıkta buhar varlığında %2-5 formaldehit çözeltisinin püskürtülmesini içerir.
Antiseptikler.
Antiseptikler, mikrobiyoloji laboratuvarlarında, öncelikle elleri yıkamak için kullanılan maddelerde bulunabilir. Tıbbi personelin antibakteriyel madde içeren maddeler kullanan hastalara acil bakım sağladığı durumlarda, bu durum hastane enfeksiyonlarını azaltır. El yıkamalarında bulunan en yaygın kimyasallar alkoller, klorheksidin glukonat, iyodoforlar, kloroksilenol ve triklosandır.
Atık ve çöpleri işlemenin geleneksel yöntemleri yakma ve buharla sterilizasyondur.
yanan atık ve enkaz işlemek için tercih edilen yöntemdir. Bu yöntem, atığı bulaşıcı olmayan hale getirir ve ayrıca şeklini ve boyutunu değiştirir. Sterilizasyon etkili bir atık arıtma yöntemidir ancak şeklini değiştirmez. 121°C'de en az 15 dakika süreyle otoklavda buharla sterilizasyon, çok sayıda bakteri sporu da dahil olmak üzere tüm mikrobiyal yaşam biçimlerini öldürür. Bu tip tam sterilizasyon 160-170ºC'de 2-4 saat kuru ısı kullanılarak da yapılabilir. Ancak steril edilecek malzeme ile kuru ısının temas etmesi sağlanmalıdır. Bu nedenle, sıvı içeren şişelerin kapakları mantar veya pamuklu çubuklarla gevşek bir şekilde kapatılmalıdır, böylece şişelerdeki hava ile buhar ve ısı alışverişi yapılabilir. Atık içeren biyolojik tehlike kapları sıkıca bağlanmalıdır. Sterilize edilmiş biyolojik tehlike materyali uygun etiketli kaplarda kapatılmalıdır.
Buhar sterilizasyonu (otoklav). Bitkisel bakteri, mantar, mikobakteri ve lipid içeren virüslerin sayısı 6 lg, bakteriyel endosporların sayısı 4 lg azaldığında bulaşıcı çöpler dekontamine edilmiş kabul edilir.
Bakterilerin beslenmesi.
Gıda. Bir bakteri hücresinin beslenmesi altında, dönüştürücü reaksiyonlar sonucunda plastik malzeme ve enerjinin absorpsiyon ve asimilasyon sürecini anlamak gerekir. . Prokaryotların beslenme türleri karmaşık ve çeşitlidir. Besinlerin bakteri hücresine girme şekline, karbon ve nitrojen kaynaklarına, enerjinin elde edilme şekline ve elektron donörlerinin doğasına bağlı olarak farklılık gösterirler.
besin maddelerinin hücre içine taşınması 3 mekanizma ile gerçekleştirilebilir: pasif difüzyon, kolaylaştırılmış difüzyon ve aktif taşıma.
pasif difüzyon maddelerin konsantrasyon gradyanı boyunca gerçekleştirilen spesifik olmayan enerjiye bağlı bir süreçtir (osmoz yasalarına göre pasif olarak daha yüksek konsantrasyonlu bir ortamdan bir madde, daha düşük konsantrasyonlu bir ortama girer). Pasif difüzyonla, bakteri hücresine, bazı iyonlara, monosakkaritlere sınırlı miktarda madde girer. Pasif difüzyon sırasında maddelerin transfer hızı önemsizdir ve taşınan moleküllerin lipofilitesine ve boyutuna bağlıdır.
Kolaylaştırılmış difüzyon permeaz enzimlerinin yardımıyla bir konsantrasyon gradyanı boyunca maddelerin uçucu olmayan bir şekilde taşınmasıdır. Permeazlar, maddelerin sitoplazmik zardan geçişini kolaylaştıran spesifik zar proteinleridir. Geçirgenlik, kendisine aktarılan maddenin bir molekülünü sabitler, bununla birlikte sitoplazmik zarın üstesinden gelir, ardından “madde-geçirgenlik” kompleksi ayrılır. Serbest kalan permeaz, diğer molekülleri iletmek için kullanılır. Prokaryotlarda hücreye sadece gliserol kolaylaştırılmış difüzyonla girer. Bu durumda, hücre içi gliserol konsantrasyonu, hücre dışındaki konsantrasyona karşılık gelir. Kolaylaştırılmış difüzyon, ökaryotik mikroorganizmaların en karakteristik özelliğidir.
aktif taşımacılık- bu, spesifik enzimlerin yardımıyla bir konsantrasyon gradyanına karşı maddelerin hücreye enerjiye bağlı transferidir. Maddelerin büyük çoğunluğu (iyonlar, karbonhidratlar, amino asitler, lipitler vb.) bakteri hücresine aktif taşıma ile girer. Aktif taşıma şu durumlarda gerçekleşebilir: (1) taşınan maddede kimyasal değişiklik olmaksızın; (2) kimyasal modifikasyon ile.
Bakteriler, hücresel yapının prokaryotik mikroorganizmalarıdır. Boyutları 0.1 ila 30 mikron arasındadır. Mikroplar son derece yaygındır. Toprakta, havada, suda, karda ve hatta kaplıcalarda, hayvanların vücudunda ve insan vücudu da dahil olmak üzere canlı organizmaların içinde yaşarlar.
Bakterilerin türlere dağılımı, mikroorganizmaların şeklinin ve mekansal dağılımının en sık dikkate alındığı çeşitli kriterlere dayanmaktadır. Böylece, hücrelerin şekline göre bakteriler ayrılır:
Coci - mikro-, diplo-, strepto-, stafilokoklar ve ayrıca sarsinler;
Çubuk şeklindeki - monobakteriler, diplobakteriler ve streptobakteriler;
Kıvrılmış türler - vibriolar ve spiroketler.
Bergey'in determinantı, bilinen tüm bakterileri, hücre duvarının yapısındaki farklılıklara ve Gram boyamaya bağlı olarak, pratik bakteriyolojide en geniş dağılımı bulan bakterilerin tanımlanması ilkelerine göre sistematize eder. Bakterilerin tanımı, familyaları, cinsleri ve türleri içeren gruplar (bölümler) tarafından verilmektedir; bazı durumlarda gruplar, sınıfları ve düzenleri içerir. İnsanlar için patojenik olan bakteriler az sayıda gruba dahildir.
Anahtar, dört ana bakteri kategorisini ayırt eder -
Gracillicutes [lat. gracilis, zarif, ince, + cutis, cilt] - ince hücre duvarına sahip türler, lekeli gram negatif;
Firmicutes [lat. flrmus, güçlü, + cutis, cilt] - kalın hücre duvarlı bakteri, lekelenme gram pozitif;
Tenericutes [lat. daha yumuşak, nazik, + cutis, cilt] - hücre duvarı olmayan bakteri(mikoplazmalar ve Mollicutes sınıfının diğer üyeleri)
Mendosicutes [lat. mendosus, düzensiz, + cutis, cilt] - arkebakteriler (metan ve sülfat indirgeyen, halofilik, termofilik ve hücre duvarından yoksun arkebakteriler).
Grup 2 Burgey'nin determinantı. Aerobik ve mikroaerofilik hareketli kıvrımlı ve kıvrımlı Gram negatif bakteriler. İnsanlar için patojenik türler, Campylobacter, Helicobacters Spirillum cinsine dahildir.
Bergey determinantının 3. grubu. Hareketsiz (nadiren hareketli) Gram negatif bakteriler. Patojen türler içermez.
Burgey determinantının 4. grubu. Gram negatif aerobik ve mikroaerofilik çubuklar ve koklar. İnsanlar için patojenik türler Legionellaceae, Neisseriaceae ve Pseudomonada-ceae familyalarına dahildir, grup ayrıca Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Francisella, Kingella ve Moraxella cinslerinin patojenik ve fırsatçı bakterilerini içerir.
Bergey determinantının 5. grubu. Fakültatif anaerobik Gram negatif çubuklar. Grup, her biri patojenik türlerin yanı sıra Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella ve Streptobacillus cinsinin patojenik ve fırsatçı bakterilerini içeren Enterobacteriaceae, Vibrionaceae ve Pasteurellaceae olmak üzere üç aileden oluşur.
Bergey determinantının 6. grubu. Gram negatif anaerobik düz, eğri ve spiral bakteriler. Patojenik ve fırsatçı türler Bacteroides, Fusobacterium, Porphoromonas ve Prevotelta cinslerine dahildir.
Bergey determinantının 7. grubu. Sülfat veya kükürt disimilasyon indirgemesi gerçekleştiren bakteri Patojenik türler içermez.
Bergey determinantının 8. grubu. Anaerobik Gram negatif koklar. Veillonella cinsinin fırsatçı bakterilerini içerir.
Bergey determinantının 9. grubu. Rickettsia ve klamidya. Üç aile - Rickettsiaceae, Bartonellaceae ve Chlamydiaceae, her biri insanlar için patojenik türler içerir.
Burgey kılavuzunun 10. ve 11. grupları, insanlar için patojenik olmayan anoksi ve oksijenli fototrofik bakterileri içerir.
Burgey determinantının 12. grubu. Aerobik kemolitotrofik bakteriler ve ilgili organizmalar. İnsanlara zarar vermeyen kükürt-demir ve manganez oksitleyici ve nitrifiye edici bakterileri birleştirir.
Burgey kılavuzunun 13 ve 14. grupları, tomurcuklanan ve/veya aşırı büyüyen bakterileri ve kılıf oluşturan bakterileri içerir. İnsanlar için patojenik olmayan, serbest yaşayan türler tarafından temsil edilir;
Burgey rehberinin 15. ve 16. grupları, meyve veren cisimler oluşturmayan kayan bakterileri birleştirir ve oluşturur. Gruplar, insanlar için patojenik türler içermez.
Burgey determinantının 17. grubu. Gram pozitif koklar. Enterococcus Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus cinslerinin fırsatçı türlerini içerir.
Burgey determinantının 18. grubu. Spor oluşturan Gram pozitif çubuklar ve koklar. Clostridium ve Bacillus cinsinin patojenik, koşullu patojenik çubuklarını içerir.
Burgey determinantının 19. grubu. Düzenli şekilli spor oluşturan Gram pozitif çubuklar. Erysipelothrix ve Listeria cinslerinin fırsatçı türleri dahil.
Burgey determinantının 20. grubu. Düzensiz şekilli spor oluşturan Gram pozitif çubuklar. Grup, Actinomyces, Corynebacterium Gardnerella, Mobiluncus, vb. cinslerinin patojenik ve fırsatçı türlerini içerir.
Burgey determinantının 21. grubu. mikobakteriler. Patojenik ve fırsatçı türleri birleştiren tek Mycobacterium cinsini içerir.
Gruplar 22-29. Aktinomisetler. Çok sayıda tür arasında sadece Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi ve Terrabacter cinslerinin nokardiyoform aktinomisetleri (Grup 22) insanlarda lezyonlara neden olabilir.
Burgey determinantının 30. grubu. Mikoplazmalar. Acholeplasma, Mycoplasma ve Ureaplasma cinsine dahil olan türler insanlar için patojeniktir.
Bergey determinantının geri kalan grupları - metanojenik bakteriler (31), sülfat indirgeyen bakteriler (32 aşırı halofilik aerobik arkebakteriler (33), hücre duvarı olmayan arkebakteriler (34), aşırı termofiller ve hipertermofiller, metabolize eden kükürt (35) - içermez İnsanlar için patojenik türler.
Bakteriler, şu anda Dünya'da var olan en eski organizma grubudur. İlk bakteriler muhtemelen 3.5 milyar yıldan daha uzun bir süre önce ortaya çıktı ve neredeyse bir milyar yıl boyunca gezegenimizdeki tek canlı yaratıklardı. Bunlar yaban hayatının ilk temsilcileri oldukları için vücutları ilkel bir yapıya sahipti.
Zamanla yapıları daha karmaşık hale geldi, ancak bugün bile bakteriler en ilkel tek hücreli organizmalar olarak kabul ediliyor. İlginç bir şekilde, bazı bakteriler eski atalarının ilkel özelliklerini hala koruyor. Bu, sıcak kükürt kaynaklarında ve rezervuarların dibindeki anoksik siltlerde yaşayan bakterilerde görülür.
Bakterilerin çoğu renksizdir. Sadece birkaçı mor veya yeşil renklidir. Ancak birçok bakterinin kolonileri, çevreye renkli bir maddenin salınması veya hücrelerin pigmentasyonu nedeniyle parlak bir renge sahiptir.
Bakteri dünyasının kaşifi, nesneleri 160-270 kat büyüten mükemmel bir büyüteç mikroskobunu ilk kez yaratan, 17. yüzyılın Hollandalı doğa bilimci Anthony Leeuwenhoek'ti.
Bakteriler prokaryot olarak sınıflandırılır ve ayrı bir krallığa ayrılır - Bakteriler.
vücut şekli
Bakteriler çok sayıda ve çeşitli organizmalardır. Form olarak farklılık gösterirler.
| bakteri adı | bakteri şekli | bakteri resmi |
| kok | küresel | |
| basil |  | Çubuk şekilli |
| titreşim | kavisli virgül | |
| Spirillum |  | Sarmal |
| streptokoklar |  | kok zinciri |
| stafilokoklar |  | kok kümeleri |
| diplokok | Bir sümüksü kapsül içine alınmış iki yuvarlak bakteri |
ulaşım yolları
Bakteriler arasında hareketli ve hareketsiz formlar vardır. Hareketli olanlar, dalga benzeri kasılmalar yoluyla veya özel bir flagellin proteininden oluşan kamçı (bükülmüş sarmal iplikler) yardımıyla hareket ederler. Bir veya daha fazla flagella olabilir. Bazı bakterilerde hücrenin bir ucunda, diğerlerinde iki veya tüm yüzey üzerinde bulunurlar.

Ancak hareket, kamçıya sahip olmayan diğer birçok bakteride de vardır. Böylece dışı mukusla kaplı olan bakteriler kayma hareketi yapabilir.
Flagella içermeyen bazı su ve toprak bakterilerinin sitoplazmasında gaz vakuolleri bulunur. Bir hücrede 40-60 koful bulunabilir. Her biri gazla doldurulur (muhtemelen azot). Vakuollerdeki gaz miktarını düzenleyerek, sudaki bakteriler su sütununa batabilir veya yüzeyine çıkabilirken, toprak bakterileri toprak kılcal damarlarında hareket edebilir.
Doğal ortam
Organizasyonun basitliği ve iddiasızlığı nedeniyle, bakteriler doğada geniş çapta dağılmıştır. Bakteriler her yerde bulunur: en saf kaynak suyunun bir damlasında bile, toprak tanelerinde, havada, kayalarda, kutup karlarında, çöl kumlarında, okyanus tabanında, çok derinlerden çıkarılan yağda ve hatta sıcak sularda. yaklaşık 80ºС sıcaklıkta kaynak suyu. Bitkilerde, meyvelerde, çeşitli hayvanlarda ve insanlarda bağırsaklarda, ağızda, uzuvlarda ve vücut yüzeyinde yaşarlar.
Bakteriler en küçük ve en çok sayıda canlılardır. Küçük boyutları nedeniyle her türlü çatlak, yarık, gözeneklere kolayca nüfuz ederler. Çok dayanıklı ve çeşitli varoluş koşullarına uyarlanmış. Canlılığını kaybetmeden kurumaya, aşırı soğuğa, 90ºº'ye kadar ısınmaya tolerans gösterirler.
Yeryüzünde bakterilerin bulunmayacağı, ancak farklı miktarlarda pratikte hiçbir yer yoktur. Bakterilerin yaşam koşulları çeşitlidir. Bazıları hava oksijenine ihtiyaç duyar, bazıları buna ihtiyaç duymaz ve oksijensiz bir ortamda yaşayabilir.
Havada: Bakteriler üst atmosfere 30 km'ye kadar yükselir. ve dahası.
Özellikle birçoğu toprakta. Bir gram toprakta yüz milyonlarca bakteri bulunabilir.
Suda: Açık rezervuarların yüzey suyu katmanlarında. Faydalı suda yaşayan bakteriler organik kalıntıları mineralize eder.
Canlı organizmalarda: patojenik bakteriler vücuda dış ortamdan girer, ancak sadece uygun koşullar altında hastalığa neden olur. Simbiyotik, sindirim organlarında yaşar, yiyecekleri parçalamaya ve özümsemeye yardımcı olur, vitaminleri sentezler.
Dış yapı
Bakteri hücresi, koruyucu ve destekleyici işlevleri yerine getiren ve ayrıca bakteriye kalıcı, karakteristik bir şekil veren hücre duvarı olan özel bir yoğun kabukla kaplanmıştır. Bir bakterinin hücre duvarı, bir bitki hücresinin kabuğuna benzer. Geçirgendir: içinden besinler hücreye serbestçe geçer ve metabolik ürünler çevreye çıkar. Bakteriler genellikle hücre duvarı üzerinde ek bir koruyucu mukus tabakası, bir kapsül geliştirir. Kapsülün kalınlığı, hücrenin çapından birçok kez daha büyük olabilir, ancak çok küçük olabilir. Kapsül hücrenin zorunlu bir parçası değildir, bakterilerin girdiği koşullara bağlı olarak oluşur. Bakterilerin kurumasını engeller.
Bazı bakterilerin yüzeyinde uzun kamçılı (bir, iki veya çok) veya kısa ince villuslar bulunur. Flagellanın uzunluğu, bakteri gövdesinin boyutundan birçok kat daha büyük olabilir. Bakteriler kamçı ve villusların yardımıyla hareket eder.

İç yapı
Bakteri hücresinin içinde yoğun bir hareketsiz sitoplazma bulunur. Katmanlı bir yapıya sahiptir, vakuol yoktur, bu nedenle sitoplazmanın özünde çeşitli proteinler (enzimler) ve yedek besinler bulunur. Bakteri hücrelerinin çekirdeği yoktur. Hücrelerinin orta kısmında kalıtsal bilgi taşıyan bir madde yoğunlaşmıştır. Bakteriler, - nükleik asit - DNA. Ancak bu madde çekirdekte çerçevelenmemiştir.
Bir bakteri hücresinin iç organizasyonu karmaşıktır ve kendine has özellikleri vardır. Sitoplazma, hücre duvarından sitoplazmik zar ile ayrılır. Sitoplazmada, ana madde veya matris, ribozomlar ve çeşitli işlevleri yerine getiren az sayıda zar yapısı (mitokondri analogları, endoplazmik retikulum, Golgi aygıtı) ayırt edilir. Bakteri hücrelerinin sitoplazması genellikle çeşitli şekil ve boyutlarda granüller içerir. Granüller, bir enerji ve karbon kaynağı olarak hizmet eden bileşiklerden oluşabilir. Bakteri hücresinde de yağ damlacıkları bulunur.
Hücrenin orta kısmında, nükleer madde olan DNA, sitoplazmadan bir zarla ayrılmadan lokalizedir. Bu çekirdeğin bir analogudur - nükleoid. Nükleoidde bir zar, çekirdekçik ve bir dizi kromozom yoktur.
Beslenme yöntemleri
Bakterilerin farklı beslenme biçimleri vardır. Bunlar arasında ototroflar ve heterotroflar bulunur. Ototroflar, beslenmeleri için bağımsız olarak organik maddeler oluşturabilen organizmalardır.
Bitkilerin nitrojene ihtiyacı vardır, ancak kendileri havadan nitrojeni ememezler. Bazı bakteriler havadaki nitrojen moleküllerini diğer moleküllerle birleştirerek bitkiler için mevcut olan maddeleri oluşturur.
Bu bakteriler genç köklerin hücrelerine yerleşir ve bu da köklerde nodül adı verilen kalınlaşmaların oluşmasına neden olur. Bu tür nodüller, baklagil ailesinin bitkilerinin ve diğer bazı bitkilerin köklerinde oluşur.

Kökler bakterilere karbonhidrat sağlar ve bakteriler köklere bitki tarafından alınabilen azot içeren maddeleri verir. İlişkileri karşılıklı olarak faydalıdır.
Bitki kökleri, bakterilerin beslendiği birçok organik madde (şekerler, amino asitler ve diğerleri) salgılar. Bu nedenle özellikle birçok bakteri kökleri çevreleyen toprak tabakasına yerleşir. Bu bakteriler, ölü bitki kalıntılarını bitkinin kullanabileceği maddelere dönüştürür. Bu toprak tabakasına rizosfer denir.
Nodül bakterilerinin kök dokulara penetrasyonu hakkında birkaç hipotez vardır:
- epidermal ve kortikal dokuya zarar vererek;
- kök kılları aracılığıyla;
- sadece genç hücre zarından;
- pektinolitik enzimler üreten eşlik eden bakteriler nedeniyle;
- Bitkilerin kök salgılarında her zaman bulunan triptofandan B-indolasetik asit sentezinin uyarılması nedeniyle.
Nodül bakterilerinin kök dokusuna giriş süreci iki aşamadan oluşur:
- kök kıllarının enfeksiyonu;
- nodül oluşum süreci.
Çoğu durumda, istilacı hücre aktif olarak çoğalır, sözde enfeksiyon ipliklerini oluşturur ve zaten bu tür iplikler şeklinde bitki dokularına hareket eder. Enfeksiyon ipliğinden ortaya çıkan nodül bakterileri konakçı dokuda çoğalmaya devam eder.

Nodül bakterilerinin hızla çoğalan hücreleriyle dolu olan bitki hücreleri yoğun bir şekilde bölünmeye başlar. Genç bir nodülün baklagil bitkisinin kökü ile bağlantısı, vasküler lifli demetler sayesinde gerçekleştirilir. İşlevsellik döneminde, nodüller genellikle yoğundur. Optimal aktivitenin tezahürü sırasında, nodüller pembe bir renk alır (legoglobin pigmenti nedeniyle). Sadece legoglobin içeren bakteriler nitrojeni sabitleyebilir.
Nodül bakterileri, toprak hektarı başına onlarca ve yüzlerce kilogram azotlu gübre oluşturur.
Metabolizma
Bakteriler metabolizmada birbirinden farklıdır. Bazıları için oksijenin katılımıyla, diğerleri için - katılımı olmadan.
Çoğu bakteri hazır organik maddelerle beslenir. Sadece birkaçı (mavi-yeşil veya siyanobakteriler) inorganik maddelerden organik maddeler oluşturabilir. Dünya atmosferinde oksijen birikmesinde önemli bir rol oynadılar.
Bakteriler dışarıdan gelen maddeleri emer, moleküllerini parçalar, kabuğunu bu parçalardan toplar ve içeriklerini yeniler (böyle büyürler) ve gereksiz molekülleri dışarı atarlar. Bakterinin kabuğu ve zarı, yalnızca doğru maddeleri emmesine izin verir.

Bakterinin kabuğu ve zarı tamamen geçirimsiz olsaydı, hücreye hiçbir madde girmezdi. Tüm maddeler için geçirgen olsaydı, hücrenin içeriği ortamla - bakterinin içinde yaşadığı çözeltiyle - karışırdı. Bakterilerin hayatta kalabilmesi için gerekli maddelerin geçmesine izin veren, ancak ihtiyaç duyulmayanların geçmesine izin vermeyen bir kabuğa ihtiyaç vardır.
Bakteri, yakınındaki besinleri emer. Sonra ne olur? Bağımsız hareket edebiliyorsa (kamçıyı hareket ettirerek veya mukusu geri iterek), gerekli maddeleri bulana kadar hareket eder.

Hareket edemiyorsa, difüzyon (bir maddenin moleküllerinin başka bir maddenin moleküllerinin kalınlığına nüfuz etme yeteneği) gerekli molekülleri kendisine getirmesini bekler.
Bakteriler, diğer mikroorganizma gruplarıyla birlikte çok büyük bir kimyasal iş yaparlar. Çeşitli bileşikleri dönüştürerek, hayati aktiviteleri için gerekli enerji ve besinleri alırlar. Metabolik süreçler, enerji elde etme yolları ve vücut maddelerini bakterilerde oluşturmak için malzeme ihtiyacı çeşitlidir.
Diğer bakteriler, vücudun organik maddelerinin sentezi için gerekli olan tüm karbon ihtiyaçlarını inorganik bileşikler pahasına karşılar. Bunlara ototrof denir. Ototrofik bakteriler, organik maddeleri inorganik olanlardan sentezleyebilir. Bunlar arasında ayırt edilir:
kemosentez
Radyan enerjinin kullanımı, karbondioksit ve sudan organik madde yaratmanın en önemli, ancak tek yolu değildir. Bakterilerin bu tür sentezler için bir enerji kaynağı olarak güneş ışığını değil, belirli inorganik bileşiklerin oksidasyonu sırasında organizmaların hücrelerinde meydana gelen kimyasal bağların enerjisini kullandığı bilinmektedir - hidrojen sülfür, kükürt, amonyak, hidrojen, nitrik asit, demir bileşikleri. demir ve manganez. Bu kimyasal enerjiyi kullanarak oluşan organik maddeyi vücutlarının hücrelerini oluşturmak için kullanırlar. Bu nedenle, bu işleme kemosentez denir.
Kemosentetik mikroorganizmaların en önemli grubu nitrifikasyon bakterileridir. Bu bakteriler toprakta yaşar ve organik kalıntıların bozunması sırasında oluşan amonyağın nitrik aside oksidasyonunu gerçekleştirir. İkincisi, toprağın mineral bileşikleri ile reaksiyona girer, nitrik asit tuzlarına dönüşür. Bu süreç iki aşamada gerçekleşir.
Demir bakterileri, demirli demiri okside dönüştürür. Oluşan demir hidroksit çöker ve sözde bataklık demir cevheri oluşturur.
Moleküler hidrojenin oksidasyonu nedeniyle bazı mikroorganizmalar bulunur, böylece ototrofik bir beslenme yolu sağlar.
Hidrojen bakterilerinin karakteristik bir özelliği, organik bileşiklerle sağlandığında ve hidrojenin yokluğunda heterotrofik bir yaşam tarzına geçme yeteneğidir.
Bu nedenle, kemoototroflar tipik ototroflardır, çünkü gerekli organik bileşikleri inorganik maddelerden bağımsız olarak sentezlerler ve bunları heterotroflar gibi diğer organizmalardan hazır olarak almazlar. Kemoototrofik bakteriler, bir enerji kaynağı olarak ışıktan tamamen bağımsız olmaları bakımından fototrofik bitkilerden farklıdır.
bakteriyel fotosentez
Spesifik pigmentler - bakteriyoklorofiller içeren bazı pigment içeren kükürt bakterileri (mor, yeşil), hidrojen sülfürün organizmalarında ayrıldığı ve ilgili bileşikleri geri yüklemek için hidrojen atomları veren güneş enerjisini emebilir. Bu sürecin fotosentez ile çok ortak noktası vardır ve yalnızca mor ve yeşil bakterilerde hidrojen sülfürün (bazen karboksilik asitler) bir hidrojen donörü olması ve yeşil bitkilerde su olması bakımından farklılık gösterir. Bunlarda ve diğerlerinde, emilen güneş ışınlarının enerjisi nedeniyle hidrojenin bölünmesi ve transferi gerçekleştirilir.
Oksijen salınımı olmadan meydana gelen bu tür bakteriyel fotosentez, fotoindirgeme olarak adlandırılır. Karbondioksitin foto-indirgenmesi, hidrojenin sudan değil, hidrojen sülfürden transferi ile ilişkilidir:
6CO 2 + 12H 2 S + hv → C6H 12 O 6 + 12S \u003d 6H 2 O
Gezegen ölçeğinde kemosentez ve bakteriyel fotosentezin biyolojik önemi nispeten küçüktür. Doğadaki kükürt döngüsünde sadece kemosentetik bakteriler önemli bir rol oynar. Yeşil bitkiler tarafından sülfürik asit tuzları şeklinde emilen kükürt restore edilir ve protein moleküllerinin bir parçasıdır. Ayrıca, ölü bitki ve hayvan kalıntılarının çürütücü bakteriler tarafından yok edilmesi sırasında, kükürt bakterileri tarafından serbest kükürte (veya sülfürik aside) oksitlenen hidrojen sülfür formunda kükürt salınır, bu da topraktaki bitkiler için mevcut sülfitleri oluşturur. Azot ve kükürt döngüsünde kemo- ve fotoototrofik bakteriler esastır.
sporlanma
Bakteri hücresinin içinde sporlar oluşur. Spor oluşumu sürecinde, bir bakteri hücresi bir dizi biyokimyasal süreçten geçer. İçindeki serbest su miktarı azalır, enzimatik aktivite azalır. Bu, sporların olumsuz çevre koşullarına (yüksek sıcaklık, yüksek tuz konsantrasyonu, kuruma vb.) karşı direncini sağlar. Spor oluşumu sadece küçük bir bakteri grubunun özelliğidir.

Sporlar, bakterilerin yaşam döngüsünde önemli bir aşama değildir. Sporülasyon, yalnızca besin eksikliği veya metabolik ürünlerin birikmesi ile başlar. Spor şeklindeki bakteriler uzun süre uykuda kalabilir. Bakteri sporları uzun süreli kaynamaya ve çok uzun süre donmaya karşı dayanıklıdır. Uygun koşullar oluştuğunda, anlaşmazlık filizlenir ve uygulanabilir hale gelir. Bakteri sporları, olumsuz koşullarda hayatta kalmaya yönelik uyarlamalardır.
üreme
Bakteriler bir hücreyi ikiye bölerek çoğalırlar. Belli bir büyüklüğe ulaşan bakteri, iki özdeş bakteriye bölünür. Sonra her biri beslenmeye, büyümeye, bölünmeye vb.

Hücrenin uzamasından sonra, yavaş yavaş enine bir septum oluşur ve ardından kızı hücreler birbirinden ayrılır; birçok bakteride, belirli koşullar altında, hücreler bölünmeden sonra karakteristik gruplar halinde bağlı kalır. Bu durumda bölme düzleminin yönüne ve bölme sayısına bağlı olarak farklı formlar ortaya çıkar. Tomurcuklanma ile üreme, istisna olarak bakterilerde meydana gelir.
Uygun koşullar altında, birçok bakteride hücre bölünmesi her 20-30 dakikada bir gerçekleşir. Bu kadar hızlı bir üreme ile 5 günde bir bakterinin yavruları tüm denizleri ve okyanusları doldurabilecek bir kütle oluşturabilmektedir. Basit bir hesap, günde 72 neslin (720.000.000.000.000.000.000.000 hücre) oluşabileceğini gösterir. Ağırlığa çevrilirse - 4720 ton. Bununla birlikte, doğada bu olmaz, çünkü çoğu bakteri güneş ışığının, kurutmanın, yiyecek eksikliğinin, 65-100ºº'ye kadar ısınmanın, türler arasındaki mücadelenin bir sonucu olarak vb. Etkisi altında hızla ölür.
Bakteri (1), yeterince besini emmiş, boyut olarak (2) büyür ve üreme (hücre bölünmesi) için hazırlanmaya başlar. DNA'sı (bir bakteride, DNA molekülü bir halkada kapalıdır) iki katına çıkar (bakteri bu molekülün bir kopyasını üretir). Her iki DNA molekülü (3.4) bakteri duvarına bağlı gibi görünmektedir ve uzadıklarında bakteriler yanlara doğru uzaklaşmaktadır (5.6). Önce nükleotid, ardından sitoplazma bölünür.
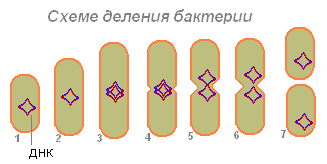
Bakteriler üzerinde iki DNA molekülünün ayrışmasından sonra, bakterinin vücudunu kademeli olarak her biri bir DNA molekülü içeren iki parçaya bölen bir daralma ortaya çıkar (7).
Olur (saman basilinde), iki bakteri birbirine yapışır ve aralarında bir köprü oluşur (1,2).

DNA, jumper (3) aracılığıyla bir bakteriden diğerine taşınır. Bir bakteride, DNA molekülleri iç içe geçer, bazı yerlerde (4) birbirine yapışır, ardından bölümleri değiştirir (5).
Bakterilerin doğadaki rolü
dolaşım
Bakteriler, doğadaki maddelerin genel dolaşımındaki en önemli halkadır. Bitkiler karbondioksit, su ve toprak mineral tuzlarından karmaşık organik maddeler oluşturur. Bu maddeler ölü mantarlar, bitkiler ve hayvan cesetleri ile toprağa geri döner. Bakteriler, karmaşık maddeleri bitkiler tarafından yeniden kullanılan basit maddelere ayrıştırır.
Bakteriler, ölü bitkilerin ve hayvan cesetlerinin karmaşık organik maddelerini, canlı organizmaların atılımlarını ve çeşitli atıkları yok eder. Bu organik maddelerle beslenen saprofitik çürüme bakterileri onları humusa dönüştürür. Bunlar gezegenimizin emirleri. Böylece bakteriler doğadaki maddelerin döngüsünde aktif olarak yer alırlar.
toprak oluşumu
Bakteriler hemen hemen her yere dağıldıkları ve çok sayıda bulundukları için, doğada meydana gelen çeşitli süreçleri büyük ölçüde belirlerler. Sonbaharda ağaçların ve çalıların yaprakları dökülür, yer üstü çim sürgünleri ölür, eski dallar dökülür ve zaman zaman yaşlı ağaçların gövdeleri düşer. Bütün bunlar yavaş yavaş humusa dönüşür. 1 cm3'te. Orman toprağının yüzey tabakası, çeşitli türden yüz milyonlarca saprofit toprak bakterisi içerir. Bu bakteriler humusu bitki kökleri tarafından topraktan emilebilen çeşitli minerallere dönüştürür.
Bazı toprak bakterileri, yaşam süreçlerinde kullanarak havadan nitrojeni emebilir. Bu nitrojen sabitleyici bakteriler kendi başlarına yaşar veya baklagil bitkilerinin köklerinde yerleşirler. Baklagillerin köklerine nüfuz eden bu bakteriler, kök hücrelerinin büyümesine ve üzerlerinde nodül oluşumuna neden olur.
Bu bakteriler, bitkilerin kullandığı azot bileşiklerini serbest bırakır. Bakteriler karbonhidratları ve mineral tuzları bitkilerden alırlar. Böylece baklagil bitkisi ile hem biri hem de diğer organizma için yararlı olan nodül bakterileri arasında yakın bir ilişki vardır. Bu fenomene simbiyoz denir.
Nodül bakterileri ile simbiyozları sayesinde baklagiller toprağı azotla zenginleştirerek verimi artırmaya yardımcı olur.
Doğada dağılım
Mikroorganizmalar her yerde bulunur. Tek istisna, aktif volkanların kraterleri ve patlatılmış atom bombalarının merkez üssündeki küçük alanlardır. Ne Antarktika'nın düşük sıcaklıkları, ne gayzerlerin kaynayan jetleri, ne tuz havuzlarındaki doymuş tuz çözeltileri, ne dağ zirvelerinin güçlü güneş ışığı, ne de nükleer reaktörlerin sert radyasyonu, mikrofloranın varlığına ve gelişimine müdahale etmez. Tüm canlılar, genellikle yalnızca depoları değil, aynı zamanda dağıtıcıları olan mikroorganizmalarla sürekli etkileşime girer. Mikroorganizmalar, aktif olarak en inanılmaz doğal substratları geliştiren gezegenimizin yerlileridir.
toprak mikroflorası
Topraktaki bakteri sayısı son derece fazladır - 1 gramda yüz milyonlarca ve milyarlarca birey. Toprakta su ve havadan çok daha fazla bulunurlar. Topraktaki toplam bakteri sayısı değişir. Bakteri sayısı toprağın türüne, durumlarına, katmanların derinliğine bağlıdır.
Mikroorganizmalar, toprak parçacıklarının yüzeyinde küçük mikrokolonilerde (her biri 20-100 hücre) bulunur. Çoğunlukla organik madde pıhtılarının kalınlığında, canlı ve ölmekte olan bitki köklerinde, ince kılcal damarlarda ve iç topaklarda gelişirler.
Toprak mikroflorası çok çeşitlidir. Burada farklı fizyolojik bakteri grupları bulunur: çürütücü, nitrifikasyon, nitrojen sabitleyici, kükürt bakterileri vb. Bunların arasında aeroblar ve anaeroblar, sporlu ve sporsuz formlar vardır. Mikroflora toprak oluşum faktörlerinden biridir.
Topraktaki mikroorganizmaların gelişme alanı, canlı bitkilerin köklerine bitişik bölgedir. Buna rizosfer denir ve içerdiği mikroorganizmaların toplamına rizosfer mikroflorası denir.
Rezervuarların mikroflorası
Su, mikroorganizmaların çok sayıda büyüdüğü doğal bir ortamdır. Çoğu suya topraktan girer. Sudaki bakteri sayısını, içindeki besin maddelerinin varlığını belirleyen bir faktör. En temizi artezyen kuyuları ve pınar sularıdır. Açık rezervuarlar ve nehirler bakteri açısından çok zengindir. En fazla sayıda bakteri, kıyıya daha yakın olan suyun yüzey katmanlarında bulunur. Kıyıdan uzaklaştıkça ve derinlik arttıkça bakteri sayısı azalır.
Saf su 1 ml'de 100-200 bakteri içerirken, kirli su 100-300 bin veya daha fazla bakteri içerir. Alt siltte, özellikle bakterilerin film oluşturduğu yüzey tabakasında çok sayıda bakteri bulunur. Bu filmde hidrojen sülfürü sülfürik aside oksitleyen ve böylece balıkların ölmesini önleyen çok sayıda kükürt ve demir bakterisi vardır. Siltte daha çok sporlu formlar bulunurken, suda sporsuz formlar baskındır.
Tür bileşimi açısından, su mikroflorası toprak mikroflorasına benzer, ancak belirli formlar da bulunur. Suya düşen çeşitli atıkları yok eden mikroorganizmalar, suyun sözde biyolojik arıtmasını yavaş yavaş gerçekleştirir.
Hava mikroflorası
Hava mikroflorası, toprak ve su mikroflorasından daha az sayıdadır. Bakteriler tozla havaya yükselir, bir süre orada kalabilir ve daha sonra yeryüzüne yerleşebilir ve beslenme yetersizliğinden veya ultraviyole ışınlarının etkisi altında ölür. Havadaki mikroorganizmaların sayısı coğrafi bölgeye, araziye, mevsime, toz kirliliğine vb. bağlıdır. Her toz zerresi bir mikroorganizma taşıyıcısıdır. Çoğu bakteri endüstriyel işletmeler üzerinde havada bulunur. Kırsaldaki hava daha temizdir. En temiz hava ormanların, dağların, karlı alanların üzerindedir. Havanın üst katmanları daha az mikrop içerir. Hava mikroflorasında, diğerlerine göre ultraviyole ışınlarına karşı daha dirençli, pigmentli ve sporlu birçok bakteri bulunur.
İnsan vücudunun mikroflorası
Bir kişinin vücudu, tamamen sağlıklı olsa bile, her zaman bir mikroflora taşıyıcısıdır. İnsan vücudu hava ve toprakla temas ettiğinde patojenler (tetanoz basili, gazlı kangren vb.) dahil olmak üzere çeşitli mikroorganizmalar giysilere ve cilde yerleşir. İnsan vücudunun açıkta kalan kısımları en sık kontamine olur. Ellerde E. coli, stafilokok bulunur. Ağız boşluğunda 100'den fazla mikrop türü vardır. Ağız, sıcaklığı, nemi, besin artıkları ile mikroorganizmaların gelişimi için mükemmel bir ortamdır.
Mide asidik bir reaksiyona sahiptir, bu nedenle içindeki mikroorganizmaların çoğu ölür. İnce bağırsaktan başlayarak reaksiyon alkali hale gelir, yani. mikroplar için uygundur. Kalın bağırsaktaki mikroflora çok çeşitlidir. Her yetişkin dışkıyla günde yaklaşık 18 milyar bakteri salgılar, yani. dünyadaki insanlardan daha fazla birey.
Dış ortama bağlı olmayan iç organlar (beyin, kalp, karaciğer, mesane vb.) genellikle mikrop içermez. Mikroplar bu organlara sadece hastalık sırasında girerler.
Bisikletteki Bakteriler
Genel olarak mikroorganizmalar ve özel olarak bakteriler, Dünya'daki maddenin biyolojik olarak önemli döngülerinde, bitkiler veya hayvanlar için tamamen erişilemeyen kimyasal dönüşümler gerçekleştirerek önemli bir rol oynar. Element döngüsünün çeşitli aşamaları, farklı türdeki organizmalar tarafından gerçekleştirilir. Her bir ayrı organizma grubunun varlığı, diğer gruplar tarafından gerçekleştirilen elementlerin kimyasal dönüşümüne bağlıdır.
nitrojen döngüsü
Azotlu bileşiklerin döngüsel dönüşümü, besin ihtiyaçları açısından çeşitli biyosfer organizmalarına gerekli azot formlarının sağlanmasında büyük bir rol oynar. Toplam nitrojen fiksasyonunun %90'ından fazlası belirli bakterilerin metabolik aktivitesinden kaynaklanmaktadır.
karbon döngüsü
Organik karbonun, moleküler oksijenin azalmasıyla birlikte karbon dioksite biyolojik dönüşümü, çeşitli mikroorganizmaların ortak metabolik aktivitesini gerektirir. Birçok aerobik bakteri, organik maddelerin tam oksidasyonunu gerçekleştirir. Aerobik koşullar altında, organik bileşikler başlangıçta fermentasyonla parçalanır ve inorganik hidrojen alıcıları (nitrat, sülfat veya CO2) varsa, fermentasyonun organik son ürünleri anaerobik solunumla daha da oksitlenir.
kükürt döngüsü
Canlı organizmalar için kükürt, esas olarak çözünür sülfatlar veya indirgenmiş organik kükürt bileşikleri şeklinde mevcuttur.
demir döngüsü
Bazı tatlı su rezervuarları yüksek konsantrasyonlarda indirgenmiş demir tuzları içerir. Bu gibi yerlerde, belirli bir bakteriyel mikroflora gelişir - indirgenmiş demiri oksitleyen demir bakterileri. Bataklık demir cevherlerinin oluşumuna ve demir tuzları bakımından zengin su kaynaklarına katılırlar.
Bakteriler, yaklaşık 3.5 milyar yıl önce Archaean'da ortaya çıkan en eski organizmalardır. Yaklaşık 2,5 milyar yıl boyunca Dünya'ya hakim oldular, biyosferi oluşturdular ve oksijen atmosferinin oluşumuna katıldılar.
Bakteriler en basit şekilde düzenlenmiş canlı organizmalardan biridir (virüsler hariç). Yeryüzünde ortaya çıkan ilk organizmalar olduklarına inanılıyor.
bakteri- Bunlar tek hücreli, klorofil içermeyen organizmalardır, vejetatif olarak bölünerek, daha az sıklıkla bağlayarak, bazen hücre içi sporlar oluşturarak çoğalırlar.
Bakterilerin boyutu mikron cinsinden ölçülür ve nadir istisnalar dışında 0,06-0,3 ile 3-5 μ arasında değişir. Bir damla su, birkaç yüz milyon bakteriyi kolayca barındırabilir.
Bir bakteri hücresinin şekli oldukça tekdüzedir. Üç ana bakteri formu bilinmektedir: yuvarlak, çubuk şeklinde ve aralarında çok sayıda ve göze çarpmayan geçişlerle kıvrımlı. Anton De Bari onları mecazi olarak bilardo topu, kurşun kalem ve tirbuşonla karşılaştırdı.
Yuvarlak şekilli bakterilere kok denir. Boyut ve göreceli konum bakımından farklılık gösterirler. Çiftler halinde bağlanan koklara diplococci, kolye şeklinde bağlanan koklara streptokoklar denir. Bölerken, karşılıklı olarak iki dik yönde dönüşümlü olarak tetracocci oluşur. Bölünme doğruysa ve birbirine dik üç yönde tekrarlanırsa, hücre bağlantıları paketler şeklinde oluşur - bunlara sardalya denir. Çok düzenli olmadan farklı yönlere bölünen koklar, üzüm fırçasını andıran rastgele kümeler oluşturur. Bunlara stafilokok denir.
Çubuk şeklindeki bakteriler görünümlerinde biraz daha çeşitlidir. Kesik veya yuvarlak uçlu, silindirik, namlu veya limon şeklinde olabilirler ve olduğu gibi ortada bir daralma ile, genellikle elipsoidal, sadece genişlikleri ve uzunlukları farklıdır. Çubuklar düz, kavisli, tekli, çiftler halinde veya zincir halinde bağlı, kısa veya çok uzun olabilir. Uzunluğu genişliğinin iki katı veya daha fazla olduğu çubuk şeklindeki bakterilere basil denir; uzunluk ve genişlik arasındaki fark küçükse, bunlara bakteri denir.
Kıvırcık şekilli bakteriler sadece uzunluk ve kalınlık bakımından değil, aynı zamanda buklelerin sayısı ve doğası bakımından da farklılık gösterir. Hafif kavisli bakterilere (kıvrılma, spiralin 1/4 turunu aşmaz) vibrio, bir veya daha fazla büyük düzenli kıvrımı olan bakterilere spirillum denir. Bazen tüm filamentin büyük eğriliklerine sahip çok sayıda küçük bukleli, kıvrımlı bir şekle sahip uzun ve ince bakterilere spiroketler denir.
Bakterilerin yapısı
Organizmalarının basitliği ve önemsiz boyutlarıyla bakteriler, en temel varlıklara aittir ve yaşamın en alt basamaklarında yer alır. Bilim ve teknolojideki muazzam ilerlemelere rağmen, bakterilerin yapısıyla ilgili tüm sorular henüz çözülmedi.
Bakteri gövdesi, hücre özü ile emprenye edilmiş, etik içerikli bir kabuk ve protoplazmadan oluşur. Bakterinin kabuğu ince, renksizdir, yapısı mikroskopta ayırt edilemez. Bunu görmek için yapay işleme yöntemlerine başvururlar. Kılıf, hücrenin dış şeklinin altında yer alır ve olumsuz koşullara karşı iyi bilinen bir savunma gibi görünmektedir. Esnekliği sayesinde hücrenin içeriğini serbestçe saran, çoğu zaman tüm vücudun hızlı hareketlerine eşlik eden bakterilerin serbest dolaşımına izin verir.
Kabuğun suyu emen dış katmanları genellikle şişer ve gözle görülür bir boyuta ulaşan jelatinimsi yapışkan bir kütle oluşturur. Dış katmanlar müsilaj hale geldikçe, kabuk protoplazmanın pahasına sürekli olarak yenilenir. Soğutulmuş yapışkan kabuğa bir kapsül denir. Mukus ve kapsül oluşumunun yoğunluğu beslenmenin özelliklerine bağlıdır ve bazen çok önemli olabilir. Mukoza kapsülü, sadece her hücrenin yakınında ayrı ayrı değil, aynı zamanda bir kolonide birbirine bağlanmış ve deyim yerindeyse ortak bir kapsül içine alınmış birçok hücrede de oluşur. Bu sümüksü bakteri kolonilerine denir. hayvanat bahçeleri. Kapsül oluşumu, tüm bakteri türlerinin özelliği değildir.
bakteri hareketi
Kendiliğinden hareket etme yeteneği, yalnızca bazı bakteri gruplarında doğaldır. Bakteriler kamçı veya kirpikler aracılığıyla hareket eder. Flagella az çok uzun filamentlere benziyor. Çok hassastırlar, incedirler, kolayca kırılırlar ve özel boyama olmadan mikroskop altında görünmezler. Çapları bakteri gövdesinin çapının 1/20'sini geçmez.
Mobil bakteri formları aşağıdaki gruplara ayrılır:
- monotrik - sadece bir polar flagellum var,
- lophotricous - hücrenin bir ucunda bir demet kamçı vardır,
- peritrik flagella vücudun tüm yüzeyinde bulunur.
Bakterinin gövdesindeki kamçının konumunun doğası, hareketinin doğasını belirler - doğrusal veya rastgele. Bakterilerin hareketliliği bir dizi koşula bağlıdır: sıcaklık, besin ortamının bileşimi, hayati aktivitelerinin ürünleri, vb. Hareketli bakteri formları, gelişimlerinin tüm aşamalarında ve tüm büyüme koşullarında değil, flagella ile donatılmaz.
sporlanma
Birçok bakterinin vücudunda, gelişimlerinin belirli dönemlerinde yuvarlak veya elipsoidal oluşumlar ortaya çıkar - destekler. Genellikle bakterilerin gelişim döngüsünü tamamlarlar. Sporların boyutu, onları üreten hücrelerin boyutuyla karşılaştırıldığında büyük ölçüde değişebilir.
Her bakteri türünde destek oluşmaz. İyi izole edilmiş, neredeyse su geçirmeyen bir kabukla çevrilidirler ve tüm canlılar dünyası arasında en kararlı oluşumlardır. Bu nedenle, genellikle birkaç saat kaynamaya ve 120 ila 140 ° arasındaki sıcaklıklarda kuru buhara uzun süre maruz kalmaya dayanırlar. Birçok basilin sporları, -190 ° sıcaklıkta ve hatta sıvı hidrojen sıcaklığında (-253 °) uzun süre kaldıktan sonra canlılıklarını korur. Ayrıca kimyasalların etkisine karşı dayanıklıdırlar - zehirler. Bütün bunlar, patojenik spor bakteri türleriyle savaşmayı son derece zorlaştırır.
Olgun bir spor, onlarca yıl canlı kalabilir. Genellikle, spor çimlenmesi belirli bir uyku döneminden sonra meydana gelir ve dış koşulların etkisiyle ilişkilidir. Tüm sporülasyon süreci bir gün veya daha kısa sürede gerçekleşir. Spor olgunlaştıktan sonra, onu üreten hücre yavaş yavaş ölür ve olgun spor dışarı çıkar. Çimlenme sırasında şişer, suda zenginleşir ve ince bir kabukla giyinmiş bir fide çıkar.
Bakterilerin üremesi
Olgunluk durumuna ve büyüme sınırına ulaşan bakteriler, basit bir bölünme ile çoğalmaya başlar. Bölünme sırasında, bakterinin gövdesinin orta kısmında bir septum belirir ve daha sonra iki yeni hücreyi böler ve ayırır. Bölünme sırasında bölümlerin sıralı düzenlenmesi farklı bakteriler için farklıdır. Çubuk şeklindeki formlarda, uzun eksene dik olarak bulunur, küresel formlarda, bölmeler bir, iki veya üç karşılıklı dik düzlemde yer alabilir, bu da streptokok, tetrakok ve sarsin gibi formların oluşumunun nedenidir. .
Bakterilerin üreme hızı bir dizi koşula bağlıdır ve çok farklı olabilir. Bakterilerin varlığı için dış koşullar ne kadar uygun olursa, bölünmeleri o kadar hızlı gerçekleşir. Normal şartlar altında bakteri sayısı yaklaşık olarak her yarım saatte bir ikiye katlanır. Her zaman engelsiz olsaydı, bir hücreden gelen bakteri sayısı devasa oranlara ulaşırdı. Mikrobiyolog Kohn'a göre, beş günde bir basilin yavruları tüm denizleri ve okyanusları doldurabilir. Ancak bu hiçbir zaman olmadı ve olmayacak. Bakterilerin yaşam döngüsü, ötesinde üremenin yavaşladığı veya tamamen durduğu belirli dış koşullarla sınırlıdır. Beslenme eksikliği, zararlı metabolik ürünler, çeşitli türlerin rekabeti vb. bakteriler üzerinde zararlı bir etkiye sahiptir. Olumsuz koşullar altında topluca ölürler.
Bakterilerin sınıflandırılması
Bakterilerin canlılar sistemindeki konumu henüz tam olarak tanımlanmamıştır. Bakterilerin bitki dünyasının bir parçası olduğu genel olarak kabul edilir ve mantarlar ve algler onlarla en yakın ilişkili organizmalardır. Çoğu durumda bakterilerin morfolojik özellikleri birkaç formla sınırlıdır: küresel, çubuklar, spiraller. Dış organizasyonlarının olağanüstü basitliği ve temel doğası, onları sınıflandırmayı zorlaştırır. Bir bakterinin türünü yalnızca morfolojik karakterlere dayanarak belirlemek imkansızdır. Bilimsel sistematiği morfolojiye ve gelişim tarihine dayanır, ancak bakterileri sınıflandırmak için sadece morfolojiyi değil, aynı zamanda fizyolojik ve biyokimyasal özelliklerini de bilmek gerekir. Bu bağlamda, bakterilerin oksijene oranı, sıcaklık koşulları, pigment oluşumu, jelatinin sıvılaşması, şekerler üzerinde asit ve gaz oluşumu, içinde bakteri üremesi ile sütün değişmesi, indol, hidrojen sülfür, amonyak oluşumu, nitratların nitritlere veya serbest nitrojene indirgenmesi. Bununla birlikte, bu her zaman bakteri türünü belirlemek için yeterli değildir.
Bakteriler için çeşitli sınıflandırma sistemleri vardır, ancak bunların hepsi koşulludur ve doğal sınıflandırmadan aşağı yukarı çok farklıdır. Fitopatojenik bakterilere uygulandığında bile bu sistemlerin veya bu durumda en az birinin dikkate alınması gerekli değildir. Sadece şu anda hemen hemen tüm fitopatojenik bakterilerin Pseudomonas, Xanthomonas, Bacterium ve Erwinia cinslerinde birleştiği söylenmelidir.
Son zamanlarda, M. V. Gorlenko (1961), Eubacteriales sınıfının fitopatojenik bakterileri için aşağıdaki sınıflandırma sistemini önerdi:
BEN. Aile Mycobacteriaceae(Chester, 1901) - hareketsiz bakteri (kamçısız):
- 1. cins - Gorynebacterium (Leman ve Neumann, 1896) - (gram-pozitif bakteri;
- 2. cins Aplanobacterium (Smith, 1905, Geshich, 1956) - gram negatif bakteriler.
II. Aile Pseudomonadaceae(Wilson ve diğerleri, 1917) - flagellalı bakteriler (polar):
- 1. cins - Pseudomonas (Migula, 1900) - boyanmamış ve floresan bakteriler;
- 2. cins - Xanthomonas (Dawson, 1839) - renkli kolonileri olan bakteriler.
III. Aile Bakterileri(Kon, 1872) - destek oluşturmayan peritrik kamçılı hareketli bakteriler:
- 1. cins - Bakteri (Ehrenberger, 1828) - pektinazlar ve protoektinazlar oluşturmayan boyanmamış formlar;
- 2. cins - Pectobacterium (Waldy, 1945) - pektinaz ve protopektinaz oluşturan boyanmamış formlar;
- 3. cins - Chromobacterium (Bergonzini, 1881) - renkli formlar.
IV. Aile Bacillaceae(Fischer, 1895) - hareketli bakteriler, spor oluşturan çubuklar:
- 1. cins - Bacillus (Kon, 1832) - hücreler spor oluşumu sırasında şişmez veya hafifçe şişmez;
- 2. cins - Clostridium (Praznovsky, 1880) - hücreler spor oluşumu sırasında şişer.
Yukarıdaki sistemde, şimdiye kadar genel olarak kabul edilen Erwinia cinsi atlanmıştır. Peritrik flagella ve pektolitik aktiviteye sahip bakterileri içeren özel bir Pectobacterium cinsi ondan izole edilir. Bu yeteneğe sahip olmayan fitopatojenik bakteriler, Bacterium cinsine atanır. Kendi içinde rasyonel olan bu sistem yenidir ve henüz günlük yaşama girmemiştir, bu nedenle bu çalışmada Erwinia cinsinin yerini aldığı sınıflandırmaya bağlıyız. Fitopatojenik bakterilerin bu jenerik adı hem ülkemizde hem de yurtdışında uzmanlaşmış literatürde yaygın olarak kullanılmaktadır.
Yapay besin ortamı kullanılmadan bakteri türünü belirlemek imkansızdır. Bu bağlamda, bakteri yetiştirirken çok karakteristik koloniler oluşturdukları not edilebilir. Bu durumda, bakteri türlerini yargılamak için bir görünüm kullanılabilir.
Bir hata bulursanız, lütfen bir metin parçasını vurgulayın ve tıklayın. Ctrl+Enter.
1.5. Bakterilerin Modern Sınıflandırması Modern bakteri taksonomisinde, diğer organizmaların sınıflandırılmasının da özelliği olan bir durum gelişmiştir: evrimsel gelişimin ana yönlerini ve temsilcilerinin ilişkilerini yansıtan bir filogenetik sınıflandırma sistemi oluşturmada ilerleme kaydedilmiştir. belirli taksonlar, ancak yapay fenomenler önemini korur - tipik sınıflandırmalar, mikroorganizmaların tanımlanması için daha uygundur. Şu anda, prokaryotların ayrıntılı bir evrimsel sistemi yoktur ve büyük olasılıkla, bu sorunun çözümü yakın bir gelecek meselesi değildir. Prokaryotların morfolojik, fizyolojik, biyokimyasal ve genetik organizasyon alanındaki özellikleri, daha yüksek organizmalar sisteminin inşasında kullanılan iyi gelişmiş ilkelerin onlara uygulanamaz olduğunu gösterir. Bakteri taksonomisi sorununun tarihsel yönleri üzerinde durmadan, prokaryotlar için en kabul edilebilir filogenetik sınıflandırma sisteminin, 16S rRNA'daki nükleotid dizisini eşleştirmeye dayalı bir sistem olduğu belirtilmelidir. Bu sistem, çok ciltli prokaryotlar ansiklopedisinin 2. baskısının temelini oluşturdu - ilk cildi 2001'de yayınlanan Bergey's Manual of Systematic Bacteriology (Bergey's Guide to Systematic Bacteria). 16S-rRNA'larının yapısına göre 26 filogenetik "dal" (grup); 23 "dal" öbakteri ve üçü arkebakteri ile temsil edilir. Bu filogenetik grupların büyük bir kısmının saf kültürler olarak izole edilmemiş ve bu nedenle henüz detaylı olarak çalışılmamış prokaryotik türleri içerdiği vurgulanmalıdır. Bu türlerin temsilcileri için şu anda sadece 16S rRNA nükleotid dizileri bilinmektedir. 23 öbakteri grubundan iki filogenetik grup gram-pozitif bakteriler tarafından temsil edilir, geri kalan gruplar gram-negatiftir. Gram negatif bakteriler, büyük bir Proteobacteria (Proteobacteria) grubundan ve bu tip hücre duvarına sahip 20 başka bakteri grubundan oluşur. Çoğu ökaryotun mitokondri ve kloroplastlarının 16S-rRNA bileşiminde en yakın olduğu Proteobakterilerin kısa bir açıklaması Tablo 1'de verilmiştir. 2. Proteobakteriler, morfolojik, fizyolojik ve biyokimyasal olarak çok heterojen olan bir gram negatif bakteri grubudur. Bu grubun temsilcileri, her türlü enerji metabolizması ve beslenme ile karakterizedir. Çoğu Proteobacteria türünün hücreleri çubuk şeklinde, küresel veya vibrioid bir şekle sahiptir; esas olarak ikili bölünme ile çoğalırlar, ancak bazı türler karmaşık bir hücre döngüsünde tomurcuklanma ve meyve veren cisimlerin oluşumu ile karakterize edilir. Bu grup hem kamçıya bağlı hareketli hem de hareketsiz bakterileri içerir. Moleküler oksijen ile ilgili olarak, Proteobakteriler zorunlu aeroblar, zorunlu ve fakültatif anaeroblar olarak sınıflandırılır. Proteobacteria grubu, 16S rRNA'daki farklılıklara dayalı olarak beş alt gruba ayrılır: alfa, beta, gama, delta ve epsilon. Proteobakterilere ek olarak, Gram-negatif öbakteriler aşağıdaki ana öbakteri gruplarını içerir: hidrojen termofilleri, yeşil filamentli bakteriler, yeşil kükürt bakterileri, siyanobakteriler, spiroketler, sitofajlar, bakteroidler, klamidya, planctomycetes, deinokoklar, kloroflexus, fusobakteriler, termofobakteriler, fibrofobakteriler vb. Filogenetik gruplar gram pozitif bakteriler - Actinobacteria ve Firmicutes. Actinobacteria grubu ("actinomycete dalı"), DNA'da yüksek GC çiftleri içeriğine sahip aşağıdaki bakteri türleri ile temsil edilir: Geodermatophilus, Frankia, Streptomyces, Arthrobacter, Micrococcus, Actinomyces, Bifidobacterium, Propionibacterium, Actinoplanes, Nocardia, Rhodococcus, Corynebacterium , Mikobakteri. Firmicutes grubu (“clostridial dal” - esas olarak DNA'da düşük GC çiftleri içeriğine sahip Gram pozitif bakteriler) aşağıdaki cinslerden oluşur: Clostridium, Lactococcus, Pediococcus, Streptococcus, Enterococcus, Leuconostoc, Listeria, Caryophanon, Staphylococcus, Sarcina , Sporosarcina, Bacillus, Desulfotomaculum, Heliobacterium, Mycoplasma, Ureaplasma, vb. Arkebakteriler üç filogenetik gruba ayrılır: Crenarchaeota, Euryarchaeota ve Korarchaeota. Crenarchaeota grubu, temsilcilerinin çoğu sülfürü metabolize eden, bazıları demir ve molibden iyonlarını azaltan aşırı derecede termofilik bakterilerden oluşur. Euryarchaeota grubu, zorunlu anaerobik metanojenik arkebakterilerin yanı sıra aşırı termofilleri ve halofilleri içerir. Korarchaeota grubu, sıcak kükürt kaynaklarında yaşayan arkebakterilerden oluşur. Bugüne kadar, bu grubun temsilcilerinin hiçbiri (benzer 16S rRNA'ya sahip) saf bir kültür olarak izole edilmedi; bu nedenle fenotipik özellikleri yeterince çalışılmamıştır. Prokaryotların filogenetik dallarının değerlendirilmesine son olarak, sadece bir ribozomal RNA geninin nükleotid dizilerinin çalışmasına dayanan önerilen filogenetik sistemin, çok sayıda sipariş vermek için teknik olarak uygun ve gelişmiş sistemlerden birinden başka bir şey olmadığı belirtilmelidir. Bu nedenle, bakterilerin doğru taksonomisini sadece bu özelliği dikkate alarak oluşturmak mantıklıdır. Bakterilerin en çok tanınan ve kullanılan fenotipik sınıflandırması, Burgey's Guide to Bacteria'nın dokuzuncu baskısında sunulan sınıflandırmadır. Bu baskıda, bakteriler hücre sınır tabakasının yapısına göre dört ana kategoriye (bölümlere) ayrılır: 1) Gracilicutes (Latin tatlılarından - deri, gracilis - ince) - hücre duvarlı gram-negatif öbakteriler; 2) Firmicutes (lat. Firmus'tan - güçlü) - hücre duvarlı gram pozitif öbakteriler; 3) Tenericutes (Latince tener - yumuşak, yumuşak) - hücre duvarlarından yoksun eubakteriler; 4) Mendosicutes (Latin mendosus'tan - hatalı) - hücre duvarları diğer prokaryotların benzer yapılarından farklı olan arkebakteriler. Gracilicutes bölümü, Gram-negatif hücre duvarına sahip çeşitli morfolojilerdeki bakterileri içerir. Üreme esas olarak ikili bölünme ile gerçekleşir, bazı bakteriler tomurcuklanma yoluyla çoğalır. Endospor oluşmaz. Çoğu hareketlidir: her türlü bakteri hareketi vardır - flagella, kayma, bükülme yardımıyla. Bölüm, aerobik, anaerobik ve fakültatif anaerobik bakterileri içerir; fototrofik ve kemotrofik bakteriler. Bölüm üç sınıfa ayrılmıştır: Scotobacteria, Oxyphotobacteria, Anoxyphotobacteria. Scotobacteria sınıfı, metabolik amaçlar için ışık enerjisini kullanmayan, ancak bunu yalnızca redoks reaksiyonlarının bir sonucu olarak alan gram negatif bakterileri içerir. Sınıfın adı Yunancadan gelmektedir. sсotos - karanlık. Bu en büyük bakteri sınıfıdır. Anoxyphotobacteria sınıfı, oksijensiz fotosentezi (moleküler oksijen salınımı olmadan) gerçekleştiren mor bakterileri, yeşil bakterileri ve heliobakterileri içerir. Oksifotobakteri sınıfı, oksijenik fotosentez (moleküler oksijen salınımı ile) gerçekleştiren siyanobakteriler ve proklorofitlerle temsil edilir. Bu tür fotosentez, bitkilerde meydana gelen fotosenteze benzer. Firmicutes bölümü, Gram pozitif hücre duvarına sahip bakterileri içerir. Hücreler farklı şekillerde olabilir: çubuklar, koklar, ipliksi, dallanma. Bazı temsilciler endosporlar oluşturur. Çoğu hareketsizdir; hareketli formlar peritrik flagellaya sahiptir. Bölüm, aerobik, anaerobik ve fakültatif anaerobik bakterileri içerir. Bölüm iki sınıftan oluşur: Firmibacteria, Thallobacteria. Firmibacteria sınıfı, çok sayıda "dallanmayan" Gram pozitif bakteri içerir. Thallobacteria sınıfı, hücreleri "dallanma" yeteneğine sahip bakterileri içerir. Tenericutes bölümü, hücre duvarı olmayan bakteriler tarafından temsil edilir. Bir hücre duvarının olmaması nedeniyle, hücrelerin şekli sabit değildir: bir türün saf kültüründe, kokoid, çubuk şeklinde, ipliksi, armut şeklinde, disk şeklinde ve diğer hücreler aynı anda bulunur. Bu bölümde yer alan bakterilerin üremesi, ikili bölünme, tomurcuklanma ile gerçekleşir. Gram boyama negatif. Agar içinde büyüyen küçük kolonilerin oluşumu karakteristiktir. Saprofitik, parazitik veya patojenik olabilirler. Bölüm, Mollicutes (mikoplazmalar) sınıfından oluşmaktadır. Mendosicutes bölümü, sert bir hücre duvarı olan ancak murein peptidoglikan içermeyen bakteriler tarafından oluşturulur. Çoğu temsilci, çoğu kamçılı olan katı anaeroblardır. Türler, ekolojik ve metabolik çeşitlilik, aşırı koşullarda yaşama yeteneği ile karakterize edilir. Bölüm bir sınıftan oluşur - Arkebakteriler. Dört bölümün (ana kategori) bir parçası olarak, sonraki bölümlerde daha fazla veya daha az ölçüde karakterize edilecek olan 35 bakteri grubu (veya bölümü) tanımlanmıştır. Aşağıdaki gruplar Gracilicutes departmanına aittir. Grup 1. Spiroketler. Grup 2. Aerobik (veya mikroaerofilik), hareketli, spiral (veya vibrioid) Gram negatif bakteriler. Grup 3. Hareketsiz veya nadiren hareketli Gram negatif kavisli bakteriler. Grup 4. Gram negatif aerobik (veya mikroaerofilik) çubuklar ve koklar. Grup 5. Opsiyonel aerobik gram negatif çubuklar. Grup 6. Gram negatif anaerobik düz, eğri veya spiral çubuklar. Grup 7. Sülfür veya sülfatın disimilasyon indirgemesini gerçekleştiren bakteriler. Grup 8. Anaerobik gram negatif koklar. Grup 9. Riketsiya ve klamidya. Grup 10. Anoksijenik fototrofik bakteriler. Grup 11. Oksijenik fototrofik bakteriler. Grup 12. Aerobik kemolitotrofik bakteriler ve ilgili organizmalar. Grup 13. Tomurcuklanma ve (veya) aşırı büyüme oluşturan bakteriler. Grup 14. Kılıflı bakteriler. Grup 15. Meyve veren cisimler oluşturmayan fotosentetik olmayan kayma bakterileri. Grup 16. Meyve veren cisimleri oluşturan kayan bakteriler. Firmicutes departmanı şunları içerir: Grup 17. Gram pozitif koklar. Grup 18. Endospor oluşturan gram pozitif çubuklar ve koklar. Grup 19. Spor oluşturmayan, doğru biçimde gram pozitif çubuklar. Grup 20. Spor oluşturmayan düzensiz şekilli Gram pozitif çubuklar. Grup 21. Mikobakteriler. Gruplar 22–29. Aktinomisetler. Tenericutes bölümüne aittir: Grup 30. Mikoplazmalar. Division Mendosicutes şunları içerir: Grup 31. Metanojenler. Grup 32. Sülfat azaltan arkebakteriler. Grup 33. Aşırı halofilik arkebakteriler (halobakteriler). Grup 34. Hücre duvarı olmayan arkebakteriler. Grup 35. Aşırı derecede termofilik ve hipertermofilik kükürt metabolize eden arkebakteriler. Sonuç olarak, doğal topluluklarda bulunan mikroorganizmaların çoğunluğunun hala saf kültürler halinde izole edilmesi gerektiği vurgulanmalıdır. Şu anda toplam mikrobiyal çeşitliliğin sadece% 0.1'inin yetiştirilebileceğine inanılıyor ve yaklaşık 5 bin prokaryot türü zaten izole edilmiş ve saf kültürde tanımlanmış olmasına rağmen, kalan bakteri temsilcilerini büyütmek ve tanımlamak mümkün değil.