Klasyfikacja mikroorganizmów według ich głównych form. Bakterie, ich różnorodność. Struktura. Witalność. Podstawowe pojęcia: bakterie
Odżywianie bakterii.
Odżywianie.
bierna dyfuzja
Ułatwiona dyfuzja
transport aktywny
W pierwszym przypadku cząsteczka składnika odżywczego tworzy kompleks z peryplazmatycznym białkiem przestrzeni, które oddziałuje ze specyficzną permeazą błony cytoplazmatycznej. Po zależnej od energii penetracji przez błonę cytoplazmatyczną kompleks „substrat – białko peryplazmatyczne – permeaza” dysocjuje z uwolnieniem cząsteczki substratu.
Podczas transportu aktywnego z chemiczną modyfikacją transportowanej substancji łańcuch zdarzeń obejmuje: (1) fosforylację enzymu błonowego-2 z cytoplazmy przez fosfoenolopirogronian; (2) wiązanie na powierzchni błony cytoplazmatycznej przez fosforylowane cząsteczki substratu enzymu-2; (3) zależny od energii transport cząsteczki substratu do cytoplazmy; (4) przeniesienie grupy fosforanowej na cząsteczkę substratu; (5) dysocjacja kompleksu „substrat-enzym” w cytoplazmie. W wyniku fosforylacji cząsteczki substratu gromadzą się w cytoplazmie komórek i nie są w stanie ich opuścić.
Klasyfikacja bakterii według rodzaju żywności.
W drodze przyjmowania składników odżywczych bakterie są podzielone na holofity oraz Holozoik. Bakterie holofitów (z greckiego. holos- kompletne i fitykos- związane z roślinami) nie są w stanie uwolnić do środowiska enzymów rozkładających substraty, w wyniku czego pobierają składniki pokarmowe wyłącznie w postaci rozpuszczonej, molekularnej. Bakterie holozoiczne (z greckiego. holos- kompletne i zoikos- spokrewnione ze zwierzętami), wręcz przeciwnie, posiadają kompleks egzoenzymów, które zapewniają odżywianie zewnętrzne - rozkład substratów do cząsteczek na zewnątrz komórki bakteryjnej. Następnie cząsteczki składników odżywczych wchodzą do wnętrza bakterii holozoicznych.
Według źródła węgla izolowany od bakterii autotrofy oraz heterotrofy. Autotrofy (z greckiego. autos- ja, trofeum- żywność) dwutlenek węgla (CO2) jest wykorzystywany jako źródło węgla, z którego syntetyzowane są wszystkie substancje zawierające węgiel. Do heterotrofy (z greckiego geteros - inny i trofeum- żywność) źródłem węgla są różne substancje organiczne w postaci cząsteczkowej (węglowodany, alkohole wielowodorotlenowe, aminokwasy, kwasy tłuszczowe). Najwyższy stopień heterotrofii jest nieodłączny od prokariontów, które mogą żyć tylko w innych żywych komórkach (na przykład rickettsia i chlamydia).
Według źródła azotu prokarionty dzieli się na 3 grupy: 1) bakterie wiążące azot (przyswajają azot cząsteczkowy z powietrza atmosferycznego); 2) bakterie zużywające nieorganiczny azot z soli amonowych, azotynów lub azotanów; 3) bakterie asymilujące azot zawarty w związkach organicznych (aminokwasy, puryny, pirymidyny itp.).
Według źródła energii bakterie dzielą się na fototrofy oraz chemotrofy. bakterie fototroficzne , podobnie jak rośliny, są w stanie wykorzystywać energię słoneczną. Fototroficzne prokariota nie powodują chorób u ludzi. Bakterie chemotroficzne otrzymywać energię w reakcjach redoks.
Z natury donorów elektronów litotrofy(z greckiego. litos- kamień) i organotrofy. Na litotrofy (chemolitotrofy ) substancje nieorganiczne działają jako donory elektronów (H 2, H 2 S, NH 3, siarka, CO, Fe 2+ itp.). Donory elektronów organotrofy (chemoorganotrofy ) to związki organiczne – węglowodany, aminokwasy itp.
Większość bakterii patogennych dla ludzi ma chemoorganotroficzny (chemoheterotroficzny) rodzaj żywienia; typ chemolitotroficzny (chemoautotroficzny) jest mniej powszechny.
Poprzez zdolność do syntezy związków organicznych Bakterie chemotroficzne dzielą się na prototrofy, auksotrofy oraz hipotrofy. bakterie prototroficzne syntetyzować wszystkie niezbędne substancje organiczne z glukozy i soli amonowych. Bakterie nazywają się auksotrofy jeśli nie są w stanie zsyntetyzować żadnej substancji organicznej ze wskazanych związków. Skrajny stopień utraty aktywności metabolicznej nazywany jest niedożywieniem. bakterie hipotroficzne zapewniają ich żywotną aktywność poprzez reorganizację struktur lub metabolitów gospodarza.
Oprócz węgla i azotu do pełnoprawnego życia bakterii niezbędne są jony siarki, fosforu i metali. Źródłami siarki są aminokwasy (cysteina, metionina), witaminy, kofaktory (biotyna, kwas liponowy itp.), siarczany. Źródłem fosforu są kwasy nukleinowe, fosfolipidy, fosforany. W wystarczająco wysokich stężeniach bakterie potrzebują magnezu, potasu, wapnia, żelaza; w znacznie mniejszych ilościach - cynk, mangan, sód, molibden, miedź, nikiel, kobalt.
czynniki wzrostowe- Są to substancje, których bakterie nie potrafią samodzielnie syntetyzować, ale bardzo ich potrzebują. Aminokwasy, zasady azotowe, witaminy, kwasy tłuszczowe, porfiryny żelaza i inne związki mogą działać jako czynniki wzrostu. Aby stworzyć optymalne warunki dla żywotnej aktywności bakterii, do pożywek należy dodać czynniki wzrostu.
Metabolizm, konwersja energii
A) Konstruktywny metabolizm.
Obowiązkową fazą odżywiania bakterii jest przyswajanie składników odżywczych, to znaczy ich włączenie w zmienionej lub zmodyfikowanej formie w reakcjach syntetycznych w celu reprodukcji składników i struktur komórkowych.
Metabolizm białek u bakterii może przebiegać w 3 fazach: pierwotny rozkład białek, wtórny rozkład białek i synteza białek. Pierwotny rozkład cząsteczek białka do peptonów jest realizowany przez egzoenzymy – egzoproteazy uwalniane przez bakterie do środowiska. Rozpad wtórny zachodzi pod wpływem endoenzymów (endoproteaz), które posiadają wszystkie bakterie. Proces ten odbywa się wewnątrz komórki bakteryjnej i polega na rozpadzie peptydów na składowe aminokwasy. Te ostatnie mogą być stosowane w postaci niezmienionej lub poddawane przemianom chemicznym (deaminacja, dekarboksylacja itp.), w wyniku których pojawiają się amoniak, indol, siarkowodór, ketokwasy, alkohol, dwutlenek węgla i inne. ma znaczenie diagnostyczne w bakteriologii.
Wraz z reakcjami rozszczepiania białek zachodzą reakcje ich syntezy. Niektóre bakterie tworzą białka z gotowych aminokwasów uzyskanych w wyniku żywienia zewnętrznego, podczas gdy inne bakterie samodzielnie syntetyzują aminokwasy z prostych związków zawierających azot i węgiel. Syntezę aminokwasów można prowadzić w reakcjach aminowania, transaminacji, amidacji, karboksylacji. Większość prokariontów jest w stanie syntetyzować wszystkie aminokwasy tworzące białka komórkowe. Cechą biosyntezy aminokwasów jest wykorzystanie powszechnych szlaków biosyntezy: cyklu kwasów trikarboksylowych, glikolizy, oksydacyjnego szlaku pentozofosforanowego itp. Głównym związkiem wyjściowym do syntezy aminokwasów jest pirogronian i fumaran.
metabolizm węglowodanów różni się autotrofami i heterotrofami (Schemat 1). Bakterie autotroficzne syntetyzują wszystkie niezbędne węglowodany z dwutlenku węgla. Surowcami do tworzenia węglowodanów w bakteriach heterotroficznych mogą być: (1) związki jedno-, dwu- i trzywęglowe; oraz (2) polisacharydy (skrobia, glikogen, celuloza). Aby podzielić te ostatnie, wiele bakterii heterotroficznych ma egzoenzymy (amylazę, pektynazę itp.), które hydrolizują polisacharydy, tworząc glukozę, maltozę, fruktozę itp.
U bakterii autotroficznych, w cyklu Calvina, z dwutlenku węgla powstaje rybulozo-fosforan-fosfor-kwas glicerynowy, który bierze udział w przebiegających w odwrotnym kierunku reakcjach glikolizy. Produktem końcowym odwróconej syntezy jest glukoza.
Bakterie heterotroficzne tworzą glukozę ze związków jedno-, dwu- i trzywęglowych, włączając je również w reakcję odwróconej glikolizy. Ze względu na to, że niektóre reakcje glikolizy są nieodwracalne, heterotrofy wytworzyły specjalne reakcje enzymatyczne, które pozwalają im ominąć nieodwracalne reakcje szlaku katabolicznego.
Gdy polisacharydy są rozszczepiane przez bakterie heterotroficzne, powstałe dwucukry wnikają do komórek i pod wpływem maltozy, sacharozy i laktozy ulegają hydrolizie i rozpadowi na monosacharydy, które następnie ulegają fermentacji lub są włączane w reakcje przekształcenia cukrów.
metabolizm lipidów. Zarówno egzogenne lipidy, jak i amfibolity metabolizmu śródmiąższowego mogą służyć jako materiały wyjściowe do tworzenia lipidów w bakteriach. Lipidy egzogenne są narażone na działanie lipaz bakteryjnych i innych enzymów lipolitycznych. Wiele rodzajów bakterii absorbuje glicerol, który służy jako źródło tworzywa sztucznego i energii. Endogennymi źródłami syntezy lipidów mogą być acetylokoenzym A, propionylo-APB, malonylo-APB (ACP - białko transportujące acetyl), fosfodioksyaceton itp.
Początkowym substratem do syntezy kwasów tłuszczowych o parzystej liczbie atomów węgla jest acetylokoenzym A, dla kwasów tłuszczowych o nieparzystej liczbie atomów węgla propionylo-APB i malonylo-APB. Tworzenie wiązań podwójnych w cząsteczce kwasu u tlenowych prokariontów zachodzi przy udziale tlenu cząsteczkowego i enzymu desaturazy. W beztlenowych prokariotach wiązania podwójne są wprowadzane na wczesnym etapie syntezy w wyniku reakcji odwodnienia. Początkowym substratem do syntezy fosfolipidów jest fosfodioksyaceton (związek pośredni szlaku glikolitycznego), którego redukcja prowadzi do powstania 3-fosforoglicerolu. Następnie do tych ostatnich dodaje się 2 reszty kwasów tłuszczowych w postaci kompleksu z APB. Produktem reakcji jest kwas fosfatydowy, którego aktywacja za pomocą CTP, a następnie przyłączenie do grupy fosforanowej seryny, inozytolu, glicerolu lub innego związku prowadzi do syntezy odpowiednich fosfolipidów.
Mikroorganizmy, które są auksotroficzne i hipotroficzne dla kwasów tłuszczowych (na przykład mykoplazmy) uzyskują je gotowe z komórek gospodarza lub pożywki.
Wymiana mononukleotydów. Mononukleotydy purynowe i pirymidynowe są niezbędnymi składnikami DNA i RNA. Wiele prokariontów potrafi zarówno wykorzystywać gotowe zasady purynowe i pirymidynowe, ich nukleozydy i nukleotydy zawarte w pożywce, jak i syntetyzować je z substancji o niskiej masie cząsteczkowej. Bakterie posiadają enzymy, które katalizują następujące etapy wzajemnej przemiany egzogennych pochodnych puryny i pirymidyny: zasada azotowa – nukleozyd – nukleotyd (mono-ditrifosforan).
Synteza mononukleotydów purynowych i pirymidynowych de novo przeprowadzane w sposób niezależny. Podczas syntezy nukleotydów purynowych w wyniku kolejnych reakcji enzymatycznych powstaje kwas inozynowy, z którego poprzez chemiczne modyfikacje pierścienia purynowego syntetyzowane są kwasy adenylowy (AMP) i guanylowy (GMP). Synteza nukleotydów pirymidynowych rozpoczyna się od powstania kwasu orotydylowego, którego dekarboksylacja daje kwas urydylowy (UMP). Z tego ostatniego powstaje UTP, którego acylacja prowadzi do powstania CTP.
Deoksyrybonukleotydy powstają w wyniku redukcji odpowiednich rybonukleotydów na poziomie difosforanów lub trifosforanów. Synteza specyficznego dla DNA nukleotydu, kwasu tymidylowego, zachodzi przez enzymatyczną metylację kwasu dezoksyurydylowego.
Wymiana jonów. Związki mineralne - jony NH 3 + , K + , Mg 2+ , Fe 2+ , SO 4 2- , PO 4 3- oraz inne bakterie pozyskiwane są ze środowiska zarówno w stanie wolnym, jak i w stanie związanym z innymi organicznymi Substancje. Kationy i aniony są transportowane do komórki bakteryjnej różnymi drogami, opisanymi w § 3. Na szybkość wnikania jonów do komórki bakteryjnej ma wpływ pH pożywki oraz aktywność fizjologiczna samych mikroorganizmów.
B) Oddychanie bakterii (metabolizm energetyczny).
Wszystkie procesy życiowe są zależne od energii, dlatego pozyskiwanie energii jest niezwykle ważnym aspektem metabolizmu prokariontów. Energię czerpią z oddychania beztlenowego i tlenowego.
Oddech, lub utlenianie biologiczne to kataboliczny proces przeniesienia elektronu z substancji donorowej na substancję akceptorową, któremu towarzyszy akumulacja energii w związkach makroergicznych . Oddychanie odbywa się w procesie reakcji katabolicznych, w wyniku których złożone substancje organiczne, rozszczepiając się, wydzielają energię i zamieniają się w proste związki. Energia zgromadzona w substancjach makroergicznych (ATP, GTP, UTP itp.) jest wykorzystywana w reakcjach anabolicznych.
Zgodnie ze sposobem oddychania mikroorganizmy dzieli się na: obligatoryjne (ścisłe) tlenowce, obligatoryjne beztlenowce oraz fakultatywne beztlenowce.
obowiązkowe aeroby potrzebujesz wolnego tlenu. Związki organiczne (węglowodany, tłuszcze, białka) są dawcami elektronów w chemoorganotroficznych tlenowcach patogennych dla ludzi, a tlen cząsteczkowy jest akceptorem elektronów. Magazynowanie energii w postaci ATP w tlenowcach chemoorganotroficznych zachodzi podczas fosforylacji oksydacyjnej donorów elektronów. Aeroby mają cytochromy (uczestniczą w przenoszeniu elektronów), a także enzymy (katalaza, dysmutaza ponadtlenkowa, peroksydaza), które inaktywują toksyczne rodniki tlenowe powstające podczas oddychania. Dysmutaza ponadtlenkowa dezaktywuje najbardziej toksyczny metabolit, rodnik ponadtlenkowy w H 2 O 2 . Katalaza enzymatyczna przekształca H 2 O 2 w H 2 O i O 2.
Specjalną grupą aerobów są bakterie mikroaerofilne, które choć potrzebują tlenu na energię, rosną lepiej przy zwiększonej zawartości CO 2 np. bakterie z rodzaju Campylobacter oraz Helicobacter.
obowiązkowe beztlenowce nie potrzebują wolnego tlenu, wręcz przeciwnie, nawet w niewielkich ilościach tlen ma na nie toksyczny wpływ. Donorami elektronów w chorobotwórczych dla człowieka beztlenowcach-chemoorganotrofach są różne związki organiczne (głównie węglowodany). Akceptorem elektronów w chemoorganotroficznych beztlenowcach są organiczne związki zawierające tlen - kwasy lub ketony, czyli akceptorem elektronów jest tlen związany z fragmentem organicznym. Magazynowanie energii w tych prokariotach następuje podczas fosforylacji substratu. Beztlenowe beztlenowce z reguły nie mają cytochromów i enzymów, które inaktywują rodniki tlenowe (ujemne wobec dysmutazy nadtlenkowej i katalazy).
W chemolitotroficznych beztlenowcach, które nie są patogenne dla ludzi, akceptorem elektronów są nieorganiczne związki zawierające tlen - azotany, siarczany, węglany.
Szczególną grupą beztlenowców są bakterie aerotolerancyjne, które mogą rosnąć w obecności tlenu atmosferycznego, ale nie wykorzystują go jako akceptora elektronów (np. bakterie kwasu mlekowego). Prokariota katalazy tolerującej powietrze i dysmutazy ponadtlenkowej są dodatnie.
Beztlenowce fakultatywne zdolny do życia zarówno w środowisku tlenowym, jak i beztlenowym. Ich donory elektronów to substancje organiczne; Akceptorami elektronów, w zależności od warunków środowiskowych, są cząsteczki lub tlen związany w związkach organicznych i nieorganicznych. Beztlenowce fakultatywne mogą akumulować energię zarówno podczas fosforylacji oksydacyjnej, jak i substratowej. Podobnie jak aeroby, ta grupa bakterii ma cytochromy i enzymy obrony antyoksydacyjnej.
Głównym substratem do pozyskiwania energii są węglowodany, które u chemoheterotroficznych prokariotów różnych typów oddychania mogą być katabolizowane do acetylokoenzymu A („aktywowany kwas octowy”). Lipidy i białka mogą pełnić rolę substratów energetycznych, ponieważ acetylokoenzym A jest również jednym z produktów pośrednich ich metabolizmu (Schemat 2).
Katabolizm węglowodanów u chemoorganotroficznych prokariontów obejmuje: (a) procesy beztlenowe — glikolizę, szlak pentozofosforanowy i szlak ketodeoksyfosfoglukonianowy; (b) proces tlenowy – cykl kwasów trikarboksylowych (cykl Krebsa). Procesy beztlenowe zachodzą u wszystkich prokariontów, podczas gdy proces tlenowy jest charakterystyczny tylko dla bezwzględnych tlenowców i fakultatywnych beztlenowców. Podstawą pozyskiwania energii drogami beztlenowymi jest fosforylacja substratu, podstawą procesu tlenowego jest fosforylacja oksydacyjna.
Definicje pojęć.
Sterylizacja, dezynfekcja i środki antyseptyczne są integralnymi elementami nowoczesnej praktyki medycznej, a zwłaszcza chirurgicznej. Zrozumienie zasad i praktyczne zastosowanie tych metod jest niezbędne, ponieważ wiele potencjalnie patogennych mikroorganizmów może przetrwać poza gospodarzem przez długi czas, wykazywać wysoką odporność na fizyczne i chemiczne środki dezynfekujące i stosunkowo łatwo przenosić się z człowieka na człowieka.
Antyseptyki- niszczenie lub zapobieganie wzrostowi drobnoustrojów chorobotwórczych lub oportunistycznych metodami chemicznymi. Termin ten jest zwykle używany w odniesieniu do zewnętrznej aplikacji preparatu chemicznego na żywe tkanki.
antyseptyczny- substancja hamująca wzrost lub niszcząca mikroorganizm (bez działania na przetrwalniki bakterii). Termin jest specyficzny dla substancji, które są stosowane do miejscowego działania na żywe tkanki.
Aseptyka oznacza brak sepsy, ale ogólnie termin ten jest używany do podkreślenia braku jakichkolwiek żywych organizmów. Metody aseptyczne oznacza każdą procedurę mającą na celu eliminację żywych organizmów i zapobieganie ich ponownemu skażeniu. Nowoczesne techniki chirurgiczne i mikrobiologiczne opierają się na procedurach aseptycznych.
Biocyd- substancja, która zabija wszystkie żywe mikroorganizmy, zarówno chorobotwórcze, jak i niepatogenne, w tym zarodniki.
Biostat- środek, który zapobiega rozwojowi drobnoustrojów, ale niekoniecznie je zabija.
Odkażenie- usuwanie drobnoustrojów bez oznaczania ilościowego. Ten termin jest względny; ostateczne usunięcie drobnoustrojów można osiągnąć poprzez sterylizację lub dezynfekcję.
Dezynfekcja- proces, który redukuje lub eliminuje wszystkie patogeny z wyjątkiem zarodników.
Środek bakteriobójczy- substancja, która niszczy mikroorganizmy, zwłaszcza patogeny. Bakteriobójczy nie niszczy zarodników.
Sanacja- metoda, dzięki której skażenie mikrobiologiczne zostaje zredukowane do „bezpiecznego” poziomu. Ta metoda była wcześniej używana do „oczyszczania” obiektów nieożywionych.
Sterylizacja- wykorzystanie czynników fizycznych i (lub) chemikaliów do całkowitego zniszczenia lub zniszczenia wszystkich form życia drobnoustrojów.
Sterylizacja.
Sterylizację definiuje się jako zniszczenie lub usunięcie (przez filtrację) wszystkich mikroorganizmów i ich zarodników. Sterylizacja jest zwykle przeprowadzana za pomocą ciepła. Sterylizacja, będąca jedną z codziennych czynności w laboratorium mikrobiologicznym, jest niezbędnym sposobem zapewnienia, że kultury, sprzęt, naczynia i pożywki wspierają wzrost tylko niezbędnych mikroorganizmów, podczas gdy inne mikroorganizmy są niszczone. Wyróżnia się takie rodzaje sterylizacji: kalcynacja w płomieniu palnika, gotowanie, działanie z przepływającą parą, para pod ciśnieniem w autoklawie, suche ciepło, pasteryzacja, tyndalizacja, chemiczna, zimna (mechaniczna) sterylizacja.
Wybór metod sterylizacji.
Przy wyborze metod sterylizacji należy wziąć pod uwagę następujące wymagania:
1. Działanie: bakteriobójcze, sporobójcze, prątkobójcze, grzybobójcze i wirusobójcze.
2. Szybkość zabiegu: sterylizację należy przeprowadzić jak najszybciej.
3. Przepuszczalność: Środki sterylizujące muszą być w stanie przeniknąć przez opakowanie i do wnętrza instrumentu.
4. Kompatybilność: nie powinno być żadnych zmian w strukturze ani funkcji materiałów, które są kilkakrotnie sterylizowane.
5. Nietoksyczność: nie powinno być zagrożenia dla zdrowia ludzkiego i środowiska.
6. Stabilność materiału organicznego: skuteczność sterylizacji nie powinna być zmniejszona w obecności materiału organicznego.
7. Adaptacyjność: możliwość zastosowania do dużych i małych ilości sterylizowanego materiału.
8. Kontrola w czasie: cykl przetwarzania musi być łatwo i dokładnie kontrolowany.
9. Cena: rozsądny koszt sprzętu, instalacji i eksploatacji.
 Fizyczne sterylizatory
Fizyczne sterylizatory
mokry serdecznie, powstający w procesie autoklawowania parowego jest głównym środkiem sterylizującym stosowanym w laboratoriach mikrobiologii klinicznej. Autoklawy służą do sterylizacji pożywek hodowlanych, materiałów ogniotrwałych i obróbki odpadów zakaźnych. Sterylizator parowy lub autoklaw to izolowana komora ciśnieniowa, która wykorzystuje nasyconą parę do generowania wysokich temperatur (rysunek 1). Powietrze jest usuwane z komory przez wypieranie masy lub podciśnienie. Najczęściej używane autoklawy z substytucją wagową. Lżejsza para jest wprowadzana do komory, aby wyprzeć cięższe powietrze. Krótka ekspozycja na parę pod ciśnieniem może zniszczyć zarodniki bakterii. W przypadku rutynowej sterylizacji pożywek hodowlanych i innych materiałów czas ekspozycji wynosi 15 minut w temperaturze 121°C i ciśnieniu 1,5 kg na 1 centymetr kwadratowy. W przypadku odpadów zakaźnych czas ekspozycji zwiększa się do 30-60 minut. Oprócz odpowiedniego czasu i temperatury bardzo ważny dla sterylizacji jest bezpośredni kontakt z parą. Podczas obchodzenia się z materiałem zakaźnym należy zapewnić maksymalne przenikanie pary do odpadów. Taki materiał musi być przetwarzany w temperaturze 132ºС. Środki przeciwnowotworowe, toksyczne chemikalia i radioizotopy, które mogą się nie rozkładać, oraz niestabilne chemikalia nie powinny być autoklawowane, ponieważ mogą parować i rozprzestrzeniać się w komorze pod wpływem ciepła.
 Sterylizacja na sucho stosowany do materiałów, które nie mogą być sterylizowane parą ze względu na możliwość uszkodzenia lub ze względu na nieprzepuszczalność materiału dla pary. Suche ciepło jest mniej skuteczne niż wilgotne ciepło i wymaga dłuższego czasu ekspozycji i wyższych temperatur. Sterylizacja suchym ciepłem jest zwykle przeprowadzana w suchej komorze grzewczej (rys. 2). Mechanizm sterylizacji suchym ciepłem jest procesem oksydacyjnym. Przykładami materiałów, w przypadku których stosuje się sterylizację na sucho, są oleje, proszki, ostre narzędzia i wyroby szklane. Jako alternatywne metody unieszkodliwiania odpadów zakaźnych stosuje się suche ciepło lub termiczną inaktywację-sterylizację.
Sterylizacja na sucho stosowany do materiałów, które nie mogą być sterylizowane parą ze względu na możliwość uszkodzenia lub ze względu na nieprzepuszczalność materiału dla pary. Suche ciepło jest mniej skuteczne niż wilgotne ciepło i wymaga dłuższego czasu ekspozycji i wyższych temperatur. Sterylizacja suchym ciepłem jest zwykle przeprowadzana w suchej komorze grzewczej (rys. 2). Mechanizm sterylizacji suchym ciepłem jest procesem oksydacyjnym. Przykładami materiałów, w przypadku których stosuje się sterylizację na sucho, są oleje, proszki, ostre narzędzia i wyroby szklane. Jako alternatywne metody unieszkodliwiania odpadów zakaźnych stosuje się suche ciepło lub termiczną inaktywację-sterylizację.
Pasteryzacja niszczy patogeny poprzez szybkie podgrzanie substancji do 71,1°C przez 15 sekund, a następnie szybkie schłodzenie. Pasteryzacja nie jest sterylizacją, ponieważ nie wszystkie mikroorganizmy są na nią wrażliwe. Metoda ta wyeliminowała przenoszenie chorób przenoszonych przez żywność, takich jak gruźlica przewodu pokarmowego i gorączka Q.
Tyndalizacja to przerywana metoda sterylizacji termicznej, którą można wykorzystać do zabicia wszystkich bakterii w roztworach. Ponieważ rosnące bakterie są łatwo zabijane przez krótkie wrzenia (5 razy w ciągu 1 godziny przez 5 minut), wystarczy odstawić roztwór na pewien czas, zanim ciepło zakłóci dojrzewanie zarodników. znaczna utrata ich odporności na ciepło.
Filtrowanie to proces, który służy do usuwania drobnoustrojów i mikroskopijnych cząstek z roztworów, powietrza i innych gazów. Najczęstszym zastosowaniem sterylizacji przez filtrację w laboratorium jest proces diagnostyczny, pożywki hodowlane, pożywki do hodowli tkankowych, surowice, roztwory zawierające składniki surowicy. Innym powszechnym zastosowaniem filtracji jest sterylizacja powietrza i gazów. Filtry membranowe z tworzywa sztucznego lub papieru, które wyróżniają się średnicą porów (od około 12 do 0,22 µm) i służą do mechanicznej separacji, służą również do zbierania drobnoustrojów z cieczy do badań mikroskopowych lub hodowli bezpośrednio na filtrze po umieszczeniu na powierzchni nasączonej odżywką średni.
promieniowanie ultrafioletowe to rodzaj promieniowania fal elektromagnetycznych, które działa na komórkowy kwas nukleinowy. Mikroorganizmy są bardzo wrażliwe na działanie promieni ultrafioletowych o długości fali 254 nm. Światło ultrafioletowe jest najczęściej używane do zabijania mikroorganizmów w powietrzu lub na powierzchniach. Inne zastosowania to sterylizacja na zimno niektórych chemikaliów i tworzyw sztucznych do zastosowań farmaceutycznych, sterylizacja surowicy do kultur komórkowych oraz dezynfekcja wody. Istotną wadą promieniowania ultrafioletowego jako sterylizatora jest jego niezdolność do penetracji materiałów.
promieniowanie jonizujące w widmie elektromagnetycznym ma zabójczy wpływ na mikroorganizmy. Widmo to obejmuje mikrofale, promienie γ, promienie rentgenowskie i przepływ elektronów. Śmiertelny efekt promieniowania jonizującego występuje w wyniku bezpośredniego działania na cząsteczkę docelową, w wyniku czego energia jest przekazywana cząsteczce; a dzięki działaniu pośredniemu - dyfuzja rodników.
energia ultradźwiękowa z niską częstotliwością dezaktywuje mikroorganizmy w roztworach wodnych. Fizyczny efekt sonikacji wynika z kawitacji. Myjki ultradźwiękowe i inne urządzenia są często używane do czyszczenia narzędzi, ale nie są uważane za sterylizatory. Jednak połączenie ultradźwięków z obróbką chemiczną zabija mikroorganizmy.
Sterylizatory chemiczne
2 % aldehyd glutarowy Jako płynny sterylizator chemiczny był wcześniej szeroko stosowany do przetwarzania materiałów medycznych i chirurgicznych, których nie można sterylizować przez ogrzewanie lub napromienianie. Aldehyd glutarowy jest również wykorzystywany do przygotowania szczepionek.
Dezynfekcja.
Dezynfekcję można przeprowadzić metodami chemicznymi lub przez gotowanie. Gotowanie to skuteczna metoda dezynfekcji narzędzi, takich jak igły i strzykawki, jeśli autoklaw nie jest dostępny. Wstępnie oczyszczone instrumenty medyczne należy gotować przez 20 minut. Dezynfekcja chemiczna jest stosowana w przypadku urządzeń wrażliwych na ciepło, które mogą zostać uszkodzone przez wysokie temperatury. Powszechnie stosowane są chemiczne środki dezynfekujące, takie jak składniki chlorowe, alkohol etylowy i izopropylowy, czwartorzędowe składniki amoniowe oraz aldehyd glutarowy.
Chemiczne środki dezynfekujące.
Alkohol (etylowy i izopropylowy), rozpuszczony w wodzie do stężenia 60-85%, jest bardzo skuteczny w dezynfekcji. Alkohole mają działanie bakteriobójcze, grzybobójcze i prątkobójcze, ale nie wpływają na zarodniki. Alkohol etylowy ma szersze spektrum działania wirusobójczego niż alkohol izopropylowy, dzięki czemu jest bardziej skuteczny wobec wirusów lipofilowych i hydrofilowych.
Rozwiązanie 37% formaldehyd, która nazywana jest formaliną, może być używana jako sterylizator, a jej stężenia 3-8% mogą być używane jako środki dezynfekujące.
Fenol w czystej postaci nie jest stosowany jako środek dezynfekujący ze względu na swoją toksyczność, zdolność do wywoływania rozwoju nowotworów i korozji. Jako środki dezynfekujące szeroko stosowane są pochodne fenolowe, w których grupa funkcyjna (chlor, brom, alkil, benzyl, fenyl, amyl) zastępuje jeden z atomów wodoru w pierścieniu aromatycznym. Ta substytucja zmniejsza wady fenolu. Składniki fenolu zabijają drobnoustroje z powodu inaktywacji układów enzymatycznych, wytrącania białek oraz rozerwania ściany komórkowej i błony komórkowej. Zwykle stosuje się stężenia 2-5%, niższe stężenia wymagają dłuższej ekspozycji.
Halogeny. W praktyce laboratoryjnej do dezynfekcji używa się wyłącznie chloru i jodu. Ze względu na fakt, że chlor jest silnym środkiem utleniającym, uważa się, że zabija drobnoustroje poprzez utlenianie. Uważa się, że jod zabija mikroorganizmy reagując z grupami N-H i S-H aminokwasów, a także z grupą fenolową aminokwasu tyrozyny i wiązaniami podwójnymi węgiel-węgiel nienasyconych kwasów tłuszczowych. Konwencjonalna obróbka polega na natryskiwaniu 2-5% roztworu formaldehydu w obecności pary o temperaturze 60-80ºC.
Antyseptyki.
Antyseptyki można znaleźć w laboratoriach mikrobiologicznych, przede wszystkim w substancjach używanych do mycia rąk. W przypadkach, gdy personel medyczny udziela pomocy doraźnej pacjentom, stosując substancje zawierające środki przeciwbakteryjne, zmniejsza to liczbę zakażeń szpitalnych. Najczęstszymi chemikaliami występującymi w płynach do mycia rąk są alkohole, glukonian chlorheksydyny, jodofory, chloroksylenol i triklosan.
Tradycyjne metody przetwarzania odpadów i śmieci to spalanie i sterylizacja parowa.
Palenie jest metodą z wyboru do obsługi odpadów i gruzu. Metoda ta sprawia, że odpady stają się niezakaźne, a także zmieniają ich kształt i wielkość. Sterylizacja jest skuteczną metodą unieszkodliwiania odpadów, ale nie zmienia jej kształtu. Sterylizacja parowa w autoklawie w temperaturze 121°C przez minimum 15 minut zabija wszystkie formy życia drobnoustrojów, w tym dużą liczbę przetrwalników bakterii. Ten rodzaj pełnej sterylizacji można również przeprowadzić przy użyciu suchego ciepła w temperaturze 160-170ºC przez 2-4 godziny. Należy jednak zapewnić, aby suche ciepło stykało się z materiałem, który ma być sterylizowany. Dlatego butelki zawierające płyn muszą być luźno zamknięte korkami lub wacikami, aby para i ciepło mogły wymieniać się z powietrzem w butelkach. Pojemniki zawierające odpady biologiczne powinny być szczelnie związane. Wysterylizowany materiał stanowiący zagrożenie biologiczne musi być zamknięty w odpowiednio oznakowanych pojemnikach.
Sterylizacja parowa (autoklaw).Śmieci zakaźne uważa się za odkażone, gdy liczba bakterii wegetatywnych, grzybów, prątków i wirusów zawierających lipidy zmniejszy się 6 lg, a przetrwalników bakteryjnych 4 lg.
Odżywianie bakterii.
Odżywianie. Pod pojęciem odżywiania komórki bakteryjnej należy rozumieć proces wchłaniania i przyswajania tworzywa sztucznego i energii w wyniku reakcji transformacyjnych . Rodzaje żywienia prokariontów są złożone i zróżnicowane. Różnią się one w zależności od sposobu, w jaki składniki odżywcze dostają się do komórki bakteryjnej, źródeł węgla i azotu, sposobu pozyskiwania energii oraz charakteru dawców elektronów.
transport składników odżywczych do komórki może być realizowany przez 3 mechanizmy: dyfuzję bierną, dyfuzję ułatwioną i transport aktywny.
bierna dyfuzja to nieswoisty proces energetycznie zależny, przebiegający wzdłuż gradientu stężeń substancji (substancja ze środowiska o wyższym stężeniu pasywnie, zgodnie z prawami osmozy, wchodzi do środowiska o niższym stężeniu). Przez dyfuzję bierną do komórki bakteryjnej dostaje się ograniczona ilość substancji, niektóre jony, monosacharydy. Szybkość przenoszenia substancji podczas dyfuzji biernej jest nieznaczna i zależy od lipofilności i wielkości transportowanych cząsteczek.
Ułatwiona dyfuzja to nielotny transport substancji wzdłuż gradientu stężeń za pomocą enzymów permeazy. Permeazy to specyficzne białka błonowe, które ułatwiają przechodzenie substancji przez błonę cytoplazmatyczną. Permeaza wiąże na sobie cząsteczkę przenoszonej substancji, wraz z którą pokonuje błonę cytoplazmatyczną, po czym kompleks „substancja-permeaza” dysocjuje. Uwolniona permeaza służy do przewodzenia innych cząsteczek. U prokariontów tylko glicerol wchodzi do komórki przez ułatwioną dyfuzję. W tym przypadku wewnątrzkomórkowe stężenie glicerolu odpowiada stężeniu na zewnątrz komórki. Ułatwiona dyfuzja jest najbardziej charakterystyczna dla mikroorganizmów eukariotycznych.
transport aktywny- jest to zależny od energii transfer substancji do komórki wbrew gradientowi stężeń za pomocą określonych enzymów. Zdecydowana większość substancji (jony, węglowodany, aminokwasy, lipidy itp.) dostaje się do komórki bakteryjnej drogą aktywnego transportu. Transport aktywny może odbywać się: (1) bez chemicznej modyfikacji transportowanej substancji; (2) z modyfikacją chemiczną.
Bakterie to mikroorganizmy prokariotyczne o strukturze komórkowej. Ich rozmiary wynoszą od 0,1 do 30 mikronów. Mikroby są niezwykle powszechne. Żyją w glebie, powietrzu, wodzie, śniegu, a nawet gorących źródłach, na ciele zwierząt, a także w organizmach żywych, w tym w organizmie człowieka.
Rozmieszczenie bakterii na gatunki opiera się na kilku kryteriach, wśród których najczęściej bierze się pod uwagę kształt drobnoustrojów oraz ich przestrzenne rozmieszczenie. Tak więc, zgodnie z kształtem komórek, bakterie dzielą się na:
Coci - mikro-, diplo-, paciorkowce-, gronkowce, a także sarcyny;
W kształcie pręcika - monobakterie, diplobakterie i paciorkowce;
Gatunki zawiłe - wibratory i krętki.
Wyznacznik Bergeya systematyzuje wszystkie znane bakterie zgodnie z zasadami identyfikacji bakterii, które w praktycznej bakteriologii znalazły najszerszą dystrybucję, opartą na różnicach w budowie ściany komórkowej iw odniesieniu do barwienia Grama. Opis bakterii jest podany w grupach (sekcjach), które obejmują rodziny, rodzaje i gatunki; w niektórych przypadkach grupy obejmują klasy i zamówienia. Bakterie chorobotwórcze dla ludzi należą do niewielkiej liczby grup.
Klucz wyróżnia cztery główne kategorie bakterii -
Gracilicutes [od łac. gracilis, wdzięczna, cienka, + cutis, skórka] - gatunek o cienkiej ścianie komórkowej, barwiony gram ujemny;
firmicutes [od łac. twarda, mocna, + skórka, skóra] - bakterie o grubej ścianie komórkowej, przebarwienia gram dodatni;
Tenericutes [od łac. Tener, Delikatny, + Cutis, Skóra] - bakterie pozbawione ściany komórkowej(mykoplazmy i inni członkowie klasy Mollicutes)
Mendosicutes [od łac. mendosus, nieregularny, + skóra skórna] - archebakterie (redukujące metan i siarczany, halofilne, termofilne i archebakterie, pozbawione ściany komórkowej).
Grupa 2 Wyznacznik Burgeya. Tlenowe i mikroaerofilne, ruchliwe, zwinięte i zakrzywione bakterie Gram-ujemne. Gatunki chorobotwórcze dla ludzi należą do rodzajów Campylobacter, Helicobacters Spirillum.
Grupa 3 wyznacznika Bergeya. Nieruchliwe (rzadko ruchliwe) bakterie Gram-ujemne. Nie zawiera gatunków chorobotwórczych.
Grupa 4 wyznacznika Burgeya. Gram-ujemne pałeczki i ziarniaki tlenowe i mikroaerofilne. Gatunki chorobotwórcze dla człowieka należą do rodzin Legionellaceae, Neisseriaceae i Pseudomonada-ceae, do grupy należą również bakterie patogenne i oportunistyczne z rodzajów Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Francisella, Kingella i Moraxella.
Grupa 5 wyznacznika Bergeya. Fakultatywne beztlenowe pałeczki Gram-ujemne. Grupę tworzą trzy rodziny - Enterobacteriaceae, Vibrionaceae i Pasteurellaceae, z których każda obejmuje gatunki chorobotwórcze, a także patogenne i oportunistyczne bakterie z rodzajów Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella i Streptobacillus.
Grupa 6 wyznacznika Bergeya. Gram-ujemne bakterie beztlenowe proste, zakrzywione i spiralne. Gatunki chorobotwórcze i oportunistyczne należą do rodzajów Bacteroides, Fusobacterium, Porphoromonas i Prevotelta.
Grupa 7 wyznacznika Bergeya. Bakterie, które przeprowadzają dysymilacyjną redukcję siarczanu lub siarki Nie obejmują gatunków patogennych.
Grupa 8 wyznacznika Bergeya. Gram-ujemne ziarniaki beztlenowe. Obejmuje bakterie oportunistyczne z rodzaju Veillonella.
Grupa 9 wyznacznika Bergeya. Rickettsia i chlamydia. Trzy rodziny - Rickettsiaceae, Bartonellaceae i Chlamydiaceae, z których każda zawiera gatunki chorobotwórcze dla człowieka.
Grupy 10 i 11 przewodnika Burgey'a obejmują anoksy- i tlenowe bakterie fototroficzne, które nie są patogenne dla ludzi.
Grupa 12 wyznacznika Burgeya. Tlenowe bakterie chemolitotroficzne i organizmy pokrewne. Łączy w sobie bakterie utleniające siarkę, żelazo i mangan oraz nitryfikujące, które nie powodują szkód u ludzi.
Grupy 13 i 14 przewodnika Burgey obejmują bakterie pączkujące i/lub rozrostowe oraz bakterie tworzące osłonki. Reprezentowane przez gatunki wolno żyjące, nie chorobotwórcze dla ludzi;
Grupy 15 i 16 przewodnika Burgey'a jednoczą bakterie szybujące, które nie tworzą owocników i je tworzą. Grupy nie obejmują gatunków chorobotwórczych dla ludzi.
Grupa 17 wyznacznika Burgeya. Cocci Gram-dodatnie. Obejmuje gatunki oportunistyczne z rodzajów Enterococcus Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus.
Grupa 18 wyznacznika Burgeya. Gram-dodatnie pałeczki i kokcy tworzące zarodniki. Obejmuje patogenne, warunkowo patogenne pałeczki z rodzajów Clostridium i Bacillus.
Grupa 19 wyznacznika Burgeya. Gram-dodatnie pałeczki tworzące zarodniki o regularnym kształcie. W tym gatunki oportunistyczne z rodzajów Erysipelothrix i Listeria.
Grupa 20 wyznacznika Burgeya. Gram-dodatnie pałeczki tworzące zarodniki o nieregularnym kształcie. Grupa obejmuje patogenne i oportunistyczne gatunki z rodzajów Actinomyces, Corynebacterium Gardnerella, Mobiluncus itp.
Grupa 21 wyznacznika Burgeya. Mykobakterie. Obejmuje jedyny rodzaj Mycobacterium, który łączy gatunki patogenne i oportunistyczne.
Grupy 22-29. Promieniowce. Spośród wielu gatunków tylko promieniowce nocardioform (grupa 22) z rodzajów Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi i Terrabacter są zdolne do wywoływania zmian u ludzi.
Grupa 30 wyznacznika Burgeya. Mykoplazmy. Gatunki należące do rodzaju Acholeplasma, Mycoplasma i Ureaplasma są patogenne dla ludzi.
Pozostałe grupy determinanty Bergeya - bakterie metanogenne (31), bakterie redukujące siarczany (32 skrajnie halofilne archebakterie tlenowe (33), archebakterie pozbawione ściany komórkowej (34), skrajnie termofile i hipertermofile, metabolizujące siarkę (35) - nie zawierają gatunki chorobotwórcze dla ludzi.
Bakterie to najstarsza grupa organizmów istniejących obecnie na Ziemi. Pierwsze bakterie pojawiły się prawdopodobnie ponad 3,5 miliarda lat temu i przez prawie miliard lat były jedynymi żywymi stworzeniami na naszej planecie. Ponieważ byli to pierwsi przedstawiciele dzikiej przyrody, ich ciało miało prymitywną strukturę.
Z biegiem czasu ich struktura stała się bardziej złożona, ale nawet dzisiaj bakterie uważane są za najbardziej prymitywne organizmy jednokomórkowe. Co ciekawe, niektóre bakterie nadal zachowują prymitywne cechy swoich starożytnych przodków. Obserwuje się to u bakterii żyjących w gorących źródłach siarkowych i anoksycznych mułach na dnie zbiorników.
Większość bakterii jest bezbarwna. Tylko kilka ma kolor fioletowy lub zielony. Ale kolonie wielu bakterii mają jasny kolor, co wynika z uwolnienia barwnej substancji do środowiska lub pigmentacji komórek.
Odkrywcą świata bakterii był Anthony Leeuwenhoek, holenderski przyrodnik z XVII wieku, który jako pierwszy stworzył doskonały mikroskop ze szkłem powiększającym, który powiększa obiekty 160-270 razy.
Bakterie są klasyfikowane jako prokariota i są podzielone na odrębne królestwo - Bakterie.
sylwetka
Bakterie to liczne i różnorodne organizmy. Różnią się formą.
| nazwa bakterii | Kształt bakterii | Obraz bakterii |
| kokcy | kulisty | |
| Bakcyl |  | w kształcie pręta |
| Wibrio | zakrzywiony przecinek | |
| Spirylla |  | Spirala |
| paciorkowce |  | Łańcuch kokcy |
| gronkowce |  | Grona kokosowe |
| diplokoki | Dwie okrągłe bakterie zamknięte w jednej oślizgłej kapsułce |
Sposoby transportu
Wśród bakterii występują formy mobilne i nieruchome. Ruchome poruszają się za pomocą skurczów falowych lub za pomocą wici (skręconych spiralnych nici), które składają się ze specjalnego białka flageliny. Może być jedna lub więcej wici. Znajdują się one u niektórych bakterii na jednym końcu komórki, u innych na dwóch lub na całej powierzchni.

Ale ruch jest również nieodłączny dla wielu innych bakterii, które nie mają wici. Tak więc bakterie pokryte śluzem na zewnątrz są zdolne do ruchu ślizgowego.
Niektóre bakterie wodne i glebowe bez wici mają wakuole gazowe w cytoplazmie. W komórce może znajdować się 40-60 wakuoli. Każdy z nich wypełniony jest gazem (przypuszczalnie azotem). Regulując ilość gazu w wakuolach, bakterie wodne mogą zatapiać się w słupie wody lub wznosić się na jej powierzchnię, podczas gdy bakterie glebowe mogą poruszać się w naczyniach włosowatych gleby.
Siedlisko
Ze względu na prostotę organizacji i bezpretensjonalność bakterie są szeroko rozpowszechnione w przyrodzie. Bakterie można znaleźć wszędzie: w kropli nawet najczystszej wody źródlanej, w ziarnach gleby, w powietrzu, na skałach, w polarnych śniegach, piaskach pustynnych, na dnie oceanu, w oleju wydobywanym z dużych głębokości, a nawet w gorącym woda źródlana o temperaturze około 80ºС. Żyją na roślinach, owocach, u różnych zwierząt i ludzi w jelitach, ustach, kończynach i na powierzchni ciała.
Bakterie to najmniejsze i najliczniejsze żywe stworzenia. Dzięki swoim niewielkim rozmiarom łatwo wnikają w wszelkie pęknięcia, szczeliny, pory. Bardzo wytrzymały i przystosowany do różnych warunków egzystencji. Tolerują suszenie, ekstremalne zimno, ogrzewanie do 90ºС, nie tracąc przy tym żywotności.
Praktycznie nie ma na Ziemi miejsca, w którym nie byłoby bakterii, ale w różnych ilościach. Warunki życia bakterii są zróżnicowane. Niektóre z nich potrzebują tlenu z powietrza, inne go nie potrzebują i są w stanie żyć w środowisku beztlenowym.
W powietrzu: bakterie wznoszą się do górnych warstw atmosfery na odległość do 30 km. i więcej.
Szczególnie dużo z nich w glebie. Jeden gram gleby może zawierać setki milionów bakterii.
W wodzie: w warstwach wód powierzchniowych zbiorników otwartych. Pożyteczne bakterie wodne mineralizują pozostałości organiczne.
W organizmach żywych: bakterie chorobotwórcze dostają się do organizmu ze środowiska zewnętrznego, ale tylko w sprzyjających warunkach powodują choroby. Symbiotyki żyją w narządach trawiennych, pomagając rozkładać i przyswajać pokarm, syntetyzują witaminy.
Struktura zewnętrzna
Komórka bakteryjna okryta jest specjalną gęstą otoczką - ścianą komórkową, która pełni funkcje ochronne i podtrzymujące, a także nadaje bakterii trwały, charakterystyczny kształt. Ściana komórkowa bakterii przypomina powłokę komórki roślinnej. Jest przepuszczalny: przez nią składniki odżywcze swobodnie przechodzą do komórki, a produkty przemiany materii trafiają do środowiska. Bakterie często tworzą dodatkową ochronną warstwę śluzu, otoczkę, na ścianie komórkowej. Grubość kapsułki może być wielokrotnie większa niż średnica samej komórki, ale może być bardzo mała. Kapsułka nie jest obowiązkową częścią komórki, powstaje w zależności od warunków, w jakich dostają się bakterie. Zapobiega wysychaniu bakterii.
Na powierzchni niektórych bakterii znajdują się długie wici (jedna, dwie lub wiele) lub krótkie cienkie kosmki. Długość wici może być wielokrotnie większa niż wielkość ciała bakterii. Bakterie poruszają się za pomocą wici i kosmków.

Struktura wewnętrzna
Wewnątrz komórki bakteryjnej znajduje się gęsta nieruchoma cytoplazma. Ma strukturę warstwową, nie ma wakuoli, więc różne białka (enzymy) i zapasowe składniki odżywcze znajdują się w samej substancji cytoplazmy. Komórki bakteryjne nie mają jądra. W centralnej części ich komórek skoncentrowana jest substancja zawierająca informacje dziedziczne. Bakterie, - kwas nukleinowy - DNA. Ale ta substancja nie jest oprawiona w jądro.
Wewnętrzna organizacja komórki bakteryjnej jest złożona i ma swoje specyficzne cechy. Cytoplazma jest oddzielona od ściany komórkowej błoną cytoplazmatyczną. W cytoplazmie wyróżnia się główną substancję lub macierz, rybosomy i niewielką liczbę struktur błonowych, które pełnią różne funkcje (analogi mitochondriów, retikulum endoplazmatyczne, aparat Golgiego). Cytoplazma komórek bakteryjnych często zawiera granulki o różnych kształtach i rozmiarach. Granulki mogą składać się ze związków, które służą jako źródło energii i węgla. Kropelki tłuszczu znajdują się również w komórce bakteryjnej.
W centralnej części komórki znajduje się substancja jądrowa, DNA, która nie jest oddzielona od cytoplazmy błoną. Jest to analog jądra - nukleoidu. Nukleoid nie posiada błony, jąderka i zestawu chromosomów.
Metody żywienia
Bakterie mają różne sposoby żywienia. Wśród nich są autotrofy i heterotrofy. Autotrofy to organizmy, które mogą samodzielnie tworzyć substancje organiczne do swojego odżywiania.
Rośliny potrzebują azotu, ale same nie mogą wchłonąć azotu z powietrza. Niektóre bakterie łączą molekuły azotu z powietrza z innymi molekułami, w wyniku czego powstają substancje dostępne dla roślin.
Bakterie te osadzają się w komórkach młodych korzeni, co prowadzi do powstawania zgrubień na korzeniach, zwanych guzkami. Takie guzki powstają na korzeniach roślin z rodziny motylkowatych i niektórych innych roślin.

Korzenie dostarczają bakteriom węglowodanów, a bakterie dostarczają korzeniom substancje zawierające azot, które mogą zostać przyswojone przez roślinę. Ich związek jest korzystny dla obu stron.
Korzenie roślin wydzielają wiele substancji organicznych (cukry, aminokwasy i inne), którymi żywią się bakterie. Dlatego szczególnie wiele bakterii osadza się w warstwie gleby otaczającej korzenie. Bakterie te przekształcają martwe resztki roślin w substancje dostępne dla rośliny. Ta warstwa gleby nazywana jest ryzosferą.
Istnieje kilka hipotez dotyczących przenikania bakterii brodawkowych do tkanek korzenia:
- poprzez uszkodzenie tkanki naskórkowej i korowej;
- przez włośniki;
- tylko przez młodą błonę komórkową;
- z powodu bakterii towarzyszących wytwarzających enzymy pektynolityczne;
- dzięki stymulacji syntezy kwasu B-indolooctowego z tryptofanu, który jest zawsze obecny w wydzielinie korzeni roślin.
Proces wprowadzania bakterii brodawkowych do tkanki korzenia składa się z dwóch faz:
- infekcja włośników;
- proces powstawania guzków.
W większości przypadków atakująca komórka aktywnie się rozmnaża, tworzy tzw. nitki infekcyjne i już w postaci takich nitek przenosi się do tkanek roślinnych. Bakterie guzkowe, które wyłoniły się z nici infekcji, nadal namnażają się w tkance gospodarza.

Wypełnione szybko namnażającymi się komórkami bakterii brodawkowych komórki roślinne zaczynają się intensywnie dzielić. Połączenie młodego guzka z korzeniem rośliny strączkowej odbywa się dzięki wiązkom naczyniowo-włóknistym. W okresie funkcjonowania guzki są zwykle gęste. Do czasu manifestacji optymalnej aktywności guzki nabierają różowego koloru (ze względu na pigment legoglobiny). Tylko te bakterie, które zawierają legoglobinę, są w stanie wiązać azot.
Bakterie brodawkowe wytwarzają dziesiątki i setki kilogramów nawozów azotowych na hektar gleby.
Metabolizm
Bakterie różnią się między sobą metabolizmem. Dla jednych idzie z udziałem tlenu, dla innych bez jego udziału.
Większość bakterii żywi się gotowymi substancjami organicznymi. Tylko nieliczne z nich (niebiesko-zielone, czyli cyjanobakterie) potrafią tworzyć substancje organiczne z nieorganicznych. Odegrały ważną rolę w akumulacji tlenu w atmosferze ziemskiej.
Bakterie wchłaniają substancje z zewnątrz, rozrywają ich molekuły, składają z tych części swoją otoczkę i uzupełniają jej zawartość (w ten sposób rosną) i wyrzucają zbędne molekuły. Otoczka i błona bakterii pozwala na wchłanianie tylko odpowiednich substancji.

Gdyby powłoka i błona bakterii były całkowicie nieprzepuszczalne, żadne substancje nie dostałyby się do komórki. Gdyby były przepuszczalne dla wszystkich substancji, zawartość komórki zmieszałaby się z pożywką - roztworem, w którym żyje bakteria. Do przetrwania bakterii potrzebna jest powłoka, która przepuszcza niezbędne substancje, ale nie te, które nie są potrzebne.
Bakteria absorbuje znajdujące się w jej pobliżu składniki odżywcze. Co się potem dzieje? Jeśli może poruszać się samodzielnie (przesuwając wić lub odpychając śluz), porusza się, aż znajdzie potrzebne substancje.

Jeśli nie może się poruszać, czeka, aż dyfuzja (zdolność cząsteczek jednej substancji do wnikania w grubość cząsteczek innej substancji) przyniesie mu niezbędne cząsteczki.
Bakterie wraz z innymi grupami mikroorganizmów wykonują ogromną chemiczną pracę. Przekształcając różne związki, otrzymują energię i składniki odżywcze niezbędne do ich życiowej aktywności. Procesy metaboliczne, sposoby pozyskiwania energii oraz zapotrzebowanie na materiały do budowy substancji ich organizmu w bakteriach są różnorodne.
Inne bakterie zaspokajają całe zapotrzebowanie na węgiel niezbędny do syntezy substancji organicznych organizmu kosztem związków nieorganicznych. Nazywane są autotrofami. Bakterie autotroficzne potrafią syntetyzować substancje organiczne z nieorganicznych. Wśród nich wyróżnia się:
Chemosynteza
Wykorzystanie energii promieniowania to najważniejszy, ale nie jedyny sposób na wytworzenie materii organicznej z dwutlenku węgla i wody. Znane są bakterie, które do takiej syntezy wykorzystują nie światło słoneczne jako źródło energii, ale energię wiązań chemicznych występujących w komórkach organizmów podczas utleniania niektórych związków nieorganicznych – siarkowodoru, siarki, amoniaku, wodoru, kwasu azotowego, związków żelaza żelazo i mangan. Wykorzystują materię organiczną utworzoną przy użyciu tej energii chemicznej do budowy komórek swojego ciała. Dlatego ten proces nazywa się chemosyntezą.
Najważniejszą grupą mikroorganizmów chemosyntetycznych są bakterie nitryfikacyjne. Bakterie te żyją w glebie i dokonują utleniania amoniaku powstałego podczas rozpadu pozostałości organicznych do kwasu azotowego. Ten ostatni, reagując ze związkami mineralnymi gleby, zamienia się w sole kwasu azotowego. Proces ten przebiega w dwóch fazach.
Bakterie żelaza przekształcają żelazo żelazawe w tlenek. Utworzony wodorotlenek żelaza osiada i tworzy tak zwaną bagienną rudę żelaza.
Niektóre mikroorganizmy istnieją w wyniku utleniania wodoru cząsteczkowego, zapewniając w ten sposób autotroficzny sposób odżywiania.
Charakterystyczną cechą bakterii wodorowych jest zdolność do przechodzenia na heterotroficzny tryb życia przy zapewnieniu związków organicznych i przy braku wodoru.
Tak więc chemoautotrofy są typowymi autotrofami, ponieważ niezależnie syntetyzują niezbędne związki organiczne z substancji nieorganicznych i nie biorą ich gotowych z innych organizmów, takich jak heterotrofy. Bakterie chemoautotroficzne różnią się od roślin fototroficznych całkowitą niezależnością od światła jako źródła energii.
fotosynteza bakteryjna
Niektóre bakterie siarkowe zawierające pigment (fioletowy, zielony), zawierające specyficzne pigmenty - bakteriochlorofile, są w stanie pochłaniać energię słoneczną, za pomocą której siarkowodór jest rozszczepiany w ich organizmach i daje atomy wodoru w celu przywrócenia odpowiednich związków. Proces ten ma wiele wspólnego z fotosyntezą i różni się jedynie tym, że u bakterii purpurowych i zielonych siarkowodorem (czasami kwasami karboksylowymi) jest dawcą wodoru, a w roślinach zielonych jest to woda. W tych i innych rozszczepianie i przenoszenie wodoru odbywa się dzięki energii zaabsorbowanych promieni słonecznych.
Taka fotosynteza bakteryjna, która zachodzi bez uwalniania tlenu, nazywana jest fotoredukcją. Fotoredukcja dwutlenku węgla wiąże się z przenoszeniem wodoru nie z wody, ale z siarkowodoru:
6CO2 + 12H2S + hv → C6H12O 6 + 12S \u003d 6H2O
Biologiczne znaczenie chemosyntezy i fotosyntezy bakteryjnej w skali planetarnej jest stosunkowo niewielkie. Tylko bakterie chemosyntetyczne odgrywają znaczącą rolę w cyklu siarkowym w przyrodzie. Wchłaniana przez rośliny zielone w postaci soli kwasu siarkowego, siarka zostaje przywrócona i staje się częścią cząsteczek białka. Ponadto, gdy martwe szczątki roślin i zwierząt są niszczone przez bakterie gnilne, siarka jest uwalniana w postaci siarkowodoru, który jest utleniany przez bakterie siarkowe do wolnej siarki (lub kwasu siarkowego), który tworzy siarczyny dostępne dla roślin w glebie. Bakterie chemo- i fotoautotroficzne są niezbędne w obiegu azotu i siarki.
zarodnikowanie
Zarodniki tworzą się wewnątrz komórki bakteryjnej. W procesie tworzenia zarodników komórka bakteryjna przechodzi szereg procesów biochemicznych. Zmniejsza się w nim ilość wolnej wody, zmniejsza się aktywność enzymatyczna. Zapewnia to odporność zarodników na niekorzystne warunki środowiskowe (wysoka temperatura, wysokie stężenie soli, wysychanie itp.). Tworzenie zarodników jest charakterystyczne tylko dla niewielkiej grupy bakterii.

Zarodniki nie są niezbędnym etapem cyklu życiowego bakterii. Zarodnikowanie zaczyna się dopiero od braku składników odżywczych lub nagromadzenia produktów przemiany materii. Bakterie w postaci zarodników mogą przez długi czas pozostawać w stanie uśpienia. Zarodniki bakterii wytrzymują długotrwałe gotowanie i bardzo długie zamrażanie. Kiedy zachodzą sprzyjające warunki, spór kiełkuje i staje się opłacalny. Zarodniki bakteryjne przystosowane są do przetrwania w niesprzyjających warunkach.
reprodukcja
Bakterie rozmnażają się, dzieląc jedną komórkę na dwie. Po osiągnięciu pewnego rozmiaru bakteria dzieli się na dwie identyczne bakterie. Potem każdy z nich zaczyna się żerować, rośnie, dzieli i tak dalej.

Po wydłużeniu komórki stopniowo tworzy się przegroda poprzeczna, a następnie komórki potomne rozchodzą się; u wielu bakterii w określonych warunkach komórki po podziale pozostają połączone w charakterystyczne grupy. W tym przypadku w zależności od kierunku płaszczyzny podziału i ilości podziałów powstają różne formy. Rozmnażanie przez pączkowanie występuje w bakteriach jako wyjątek.
W sprzyjających warunkach podział komórek u wielu bakterii następuje co 20-30 minut. Przy tak szybkim rozmnażaniu potomstwo jednej bakterii w ciągu 5 dni jest w stanie uformować masę, która może wypełnić wszystkie morza i oceany. Proste obliczenia pokazują, że dziennie mogą powstawać 72 pokolenia (72 000 000 000 000 000 000 000 komórek). W przeliczeniu na wagę - 4720 ton. Jednak tak się nie dzieje w naturze, ponieważ większość bakterii szybko umiera pod wpływem światła słonecznego, suszenia, braku jedzenia, ogrzewania do 65-100ºС w wyniku walki między gatunkami itp.
Bakteria (1), po wchłonięciu wystarczającej ilości pokarmu, powiększa się (2) i zaczyna przygotowywać się do rozmnażania (podział komórek). Jej DNA (w bakterii cząsteczka DNA jest zamknięta w pierścieniu) podwaja się (bakteria wytwarza kopię tej cząsteczki). Wydaje się, że obie cząsteczki DNA (3.4) są przyczepione do ściany bakteryjnej, a po wydłużeniu bakterie rozchodzą się na boki (5.6). Najpierw dzieli się nukleotyd, potem cytoplazma.
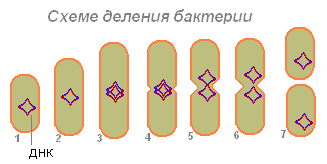
Po dywergencji dwóch cząsteczek DNA na bakteriach pojawia się zwężenie, które stopniowo dzieli organizm bakterii na dwie części, z których każda zawiera cząsteczkę DNA (7).
Zdarza się (w pałeczkach siana) dwie bakterie sklejają się i tworzy się między nimi most (1,2).

DNA jest transportowane z jednej bakterii do drugiej przez zworkę (3). W jednej bakterii cząsteczki DNA splatają się, sklejają w niektórych miejscach (4), po czym zamieniają się fragmentami (5).
Rola bakterii w przyrodzie
Krążenie
Bakterie są najważniejszym ogniwem w ogólnym obiegu substancji w przyrodzie. Rośliny tworzą złożone substancje organiczne z dwutlenku węgla, wody i soli mineralnych gleby. Substancje te wracają do gleby wraz z martwymi grzybami, roślinami i zwłokami zwierząt. Bakterie rozkładają złożone substancje na proste, które są ponownie wykorzystywane przez rośliny.
Bakterie niszczą złożoną materię organiczną martwych roślin i zwłok zwierząt, wydzieliny organizmów żywych i różne odpady. Żywiąc się tymi substancjami organicznymi, bakterie saprofityczne zamieniają je w próchnicę. To są sanitariusze naszej planety. W ten sposób bakterie są aktywnie zaangażowane w cykl substancji w przyrodzie.
tworzenie gleby
Ponieważ bakterie są rozprzestrzenione niemal wszędzie i występują w ogromnych ilościach, w dużej mierze determinują różne procesy zachodzące w przyrodzie. Jesienią opadają liście drzew i krzewów, zamierają naziemne pędy traw, odpadają stare gałęzie, a od czasu do czasu opadają pnie starych drzew. Wszystko to stopniowo zamienia się w próchnicę. W 1 cm 3. W warstwie powierzchniowej gleby leśnej znajdują się setki milionów saprofitycznych bakterii glebowych kilku gatunków. Bakterie te przekształcają próchnicę w różne minerały, które mogą być przyswajane z gleby przez korzenie roślin.
Niektóre bakterie glebowe są w stanie pobierać azot z powietrza, wykorzystując go w procesach życiowych. Te bakterie wiążące azot żyją samodzielnie lub zasiedlają korzenie roślin strączkowych. Po przeniknięciu do korzeni roślin strączkowych bakterie te powodują wzrost komórek korzeniowych i tworzenie na nich guzków.
Bakterie te uwalniają związki azotu, z których korzystają rośliny. Bakterie pozyskują z roślin węglowodany i sole mineralne. Tak więc istnieje ścisły związek między rośliną strączkową a bakteriami brodawkowymi, co jest przydatne zarówno dla jednego, jak i drugiego organizmu. Zjawisko to nazywa się symbiozą.
Dzięki symbiozie z bakteriami brodawkowymi rośliny strączkowe wzbogacają glebę w azot, przyczyniając się do zwiększenia plonów.
Dystrybucja w przyrodzie
Mikroorganizmy są wszechobecne. Jedynymi wyjątkami są kratery aktywnych wulkanów i niewielkie obszary w epicentrach zdetonowanych bomb atomowych. Ani niskie temperatury Antarktydy, ani wrzące strumienie gejzerów, ani nasycone roztwory soli w basenach solnych, ani silne nasłonecznienie górskich szczytów, ani ostre promieniowanie reaktorów jądrowych nie przeszkadzają w istnieniu i rozwojowi mikroflory. Wszystkie żywe istoty nieustannie wchodzą w interakcję z mikroorganizmami, będąc często nie tylko ich magazynami, ale także dystrybutorami. Mikroorganizmy są tubylcami naszej planety, aktywnie rozwijającymi najbardziej niesamowite naturalne podłoża.
Mikroflora glebowa
Liczba bakterii w glebie jest niezwykle duża - setki milionów i miliardy osobników w 1 gramie. Występują znacznie liczniej w glebie niż w wodzie i powietrzu. Całkowita liczba bakterii w glebie jest różna. Liczba bakterii zależy od rodzaju gleby, ich stanu, głębokości warstw.
Na powierzchni cząstek gleby mikroorganizmy znajdują się w małych mikrokoloniach (po 20-100 komórek). Często rozwijają się w grubości skrzepów materii organicznej, na żywych i obumierających korzeniach roślin, w cienkich naczyniach włosowatych i wewnątrz grudek.
Mikroflora glebowa jest bardzo zróżnicowana. Występują tu różne fizjologiczne grupy bakterii: bakterie gnilne, nitryfikacyjne, wiążące azot, siarkowe itp. wśród nich są tlenowce i beztlenowce, formy przetrwalnikowe i bezprzetrwalnikowe. Mikroflora jest jednym z czynników kształtowania gleby.
Obszar rozwoju mikroorganizmów w glebie to strefa przylegająca do korzeni żywych roślin. Nazywa się ryzosferą, a całość zawartych w niej mikroorganizmów nazywana jest mikroflorą ryzosfery.
Mikroflora zbiorników wodnych
Woda to naturalne środowisko, w którym licznie rozwijają się mikroorganizmy. Większość z nich dostaje się do wody z gleby. Czynnik, który determinuje liczbę bakterii w wodzie, obecność w niej składników odżywczych. Najczystsze są wody studni i źródeł artezyjskich. Otwarte zbiorniki i rzeki są bardzo bogate w bakterie. Najwięcej bakterii znajduje się w powierzchniowych warstwach wody, bliżej brzegu. Wraz ze wzrostem odległości od wybrzeża i głębokością zmniejsza się liczba bakterii.
Czysta woda zawiera 100-200 bakterii w 1 ml, a zanieczyszczona 100-300 tysięcy i więcej. W mule dennym znajduje się wiele bakterii, zwłaszcza w warstwie powierzchniowej, gdzie bakterie tworzą film. W tym filmie jest wiele bakterii siarkowych i żelaznych, które utleniają siarkowodór do kwasu siarkowego, zapobiegając w ten sposób umieraniu ryb. W mule występuje więcej form zarodnikowych, podczas gdy w wodzie przeważają formy bez zarodników.
Pod względem składu gatunkowego mikroflora wodna jest zbliżona do mikroflory glebowej, ale spotykane są również formy specyficzne. Niszcząc różne odpady, które dostały się do wody, mikroorganizmy stopniowo przeprowadzają tak zwane biologiczne oczyszczanie wody.
Mikroflora powietrza
Mikroflora powietrza jest mniej liczna niż mikroflora glebowa i wodna. Bakterie unoszą się w powietrze z kurzem, mogą tam przebywać przez chwilę, a następnie osiadać na powierzchni ziemi i ginąć z braku pożywienia lub pod wpływem promieni ultrafioletowych. Ilość drobnoustrojów w powietrzu zależy od obszaru geograficznego, ukształtowania terenu, pory roku, zanieczyszczenia pyłowego itp. Każda drobinka kurzu jest nośnikiem drobnoustrojów. Większość bakterii w powietrzu nad przedsiębiorstwami przemysłowymi. Powietrze na wsi jest czystsze. Najczystsze powietrze jest nad lasami, górami, zaśnieżonymi przestrzeniami. Górne warstwy powietrza zawierają mniej zarazków. W mikroflorze powietrza znajduje się wiele bakterii pigmentowych i zarodnikowych, które są bardziej odporne niż inne na promienie ultrafioletowe.
Mikroflora ludzkiego ciała
Ciało człowieka, nawet całkowicie zdrowego, jest zawsze nośnikiem mikroflory. Gdy ciało ludzkie wchodzi w kontakt z powietrzem i glebą, na odzieży i skórze osadzają się różne mikroorganizmy, w tym patogeny (pałeczki tężca, zgorzel gazowa itp.). Skażone są najczęściej odsłonięte części ludzkiego ciała. Na dłoniach znajdują się E. coli, gronkowce. W jamie ustnej występuje ponad 100 rodzajów drobnoustrojów. Usta ze swoją temperaturą, wilgotnością, resztkami składników odżywczych są doskonałym środowiskiem do rozwoju mikroorganizmów.
Żołądek ma odczyn kwaśny, więc większość mikroorganizmów w nim ginie. Począwszy od jelita cienkiego reakcja staje się zasadowa, tj. korzystny dla drobnoustrojów. Mikroflora w jelicie grubym jest bardzo zróżnicowana. Każdy dorosły wydala około 18 miliardów bakterii dziennie wraz z odchodami, tj. więcej osób niż ludzi na świecie.
Narządy wewnętrzne, które nie są połączone ze środowiskiem zewnętrznym (mózg, serce, wątroba, pęcherz itp.) są zwykle wolne od drobnoustrojów. Mikroby dostają się do tych organów tylko podczas choroby.
Bakterie w kolarstwie
Ogólnie rzecz biorąc, mikroorganizmy, aw szczególności bakterie, odgrywają ważną rolę w biologicznie ważnych cyklach materii na Ziemi, dokonując przemian chemicznych całkowicie niedostępnych ani dla roślin, ani dla zwierząt. Różne etapy cyklu pierwiastków są realizowane przez organizmy różnych typów. Istnienie każdej oddzielnej grupy organizmów zależy od chemicznej przemiany pierwiastków dokonywanej przez inne grupy.
obieg azotu
Cykliczna przemiana związków azotowych odgrywa nadrzędną rolę w dostarczaniu niezbędnych form azotu do różnych organizmów biosfery z punktu widzenia potrzeb żywieniowych. Ponad 90% całkowitego wiązania azotu wynika z aktywności metabolicznej niektórych bakterii.
Cykl węgla
Biologiczna przemiana węgla organicznego w dwutlenek węgla, połączona z redukcją tlenu cząsteczkowego, wymaga wspólnej aktywności metabolicznej różnych mikroorganizmów. Wiele bakterii tlenowych przeprowadza całkowite utlenianie substancji organicznych. W warunkach tlenowych związki organiczne są początkowo rozkładane przez fermentację, a końcowe produkty fermentacji są dalej utleniane przez oddychanie beztlenowe, jeśli obecne są nieorganiczne akceptory wodoru (azotan, siarczan lub CO2).
Cykl siarki
Dla organizmów żywych siarka jest dostępna głównie w postaci rozpuszczalnych siarczanów lub zredukowanych organicznych związków siarki.
Cykl żelaza
Niektóre zbiorniki słodkiej wody zawierają wysokie stężenia zredukowanych soli żelaza. W takich miejscach rozwija się specyficzna mikroflora bakteryjna - bakterie żelazowe, które utleniają zredukowane żelazo. Uczestniczą w tworzeniu bagiennych rud żelaza i wód bogatych w sole żelaza.
Bakterie to najstarsze organizmy, które pojawiły się około 3,5 miliarda lat temu w Archaeanie. Przez około 2,5 miliarda lat dominowali na Ziemi, tworząc biosferę i uczestniczyli w tworzeniu atmosfery tlenowej.
Bakterie to jedne z najprostszych organizmów żywych (poza wirusami). Uważa się, że są pierwszymi organizmami, które pojawiły się na Ziemi.
bakteria- Są to organizmy jednokomórkowe, wolne od chlorofilu, które rozmnażają się wegetatywnie przez podziały, rzadziej przez sploty, czasem tworząc wewnątrzkomórkowe zarodniki.
Wielkość bakterii mierzona jest w mikronach i, z rzadkimi wyjątkami, waha się od 0,06-0,3 do 3-5 μm. Kropla wody może z łatwością zawierać kilkaset milionów bakterii.
Kształt komórki bakteryjnej jest dość jednolity. Znane są trzy główne formy bakterii: okrągłe, pręcikowe i zawiłe, z licznymi i niepozornymi przejściami między nimi. Anton De Bari w przenośni porównał je z kulą bilardową, ołówkiem i korkociągiem.
Bakterie o okrągłym kształcie nazywane są cocci. Różnią się wielkością i względną pozycją. Ziarniczki połączone parami nazywane są diplokokami, ale połączone w formie naszyjnika nazywane są paciorkowcami. Podczas dzielenia, naprzemiennie w dwóch wzajemnie prostopadłych kierunkach, powstają tetrakoki. Jeśli podział jest prawidłowy i powtarza się w trzech wzajemnie prostopadłych kierunkach, to powstają połączenia komórkowe w postaci paczek – są to tzw. sardynki. Dzieląc się w różnych kierunkach bez większej regularności, ziarniaki tworzą losowe kiście przypominające pędzel winogron. Nazywane są gronkowcami.
Bakterie w kształcie pręcików mają nieco bardziej zróżnicowany wygląd. Mogą mieć końce ścięte lub zaokrąglone, cylindryczne, beczkowate lub cytrynowe i niejako ze zwężeniem pośrodku, często elipsoidalnym, różniące się jedynie szerokością i długością. Kije mogą być proste, zakrzywione, pojedyncze, połączone parami lub łańcuchem, krótkie lub mocno wydłużone. Bakterie w kształcie pręcików, których długość jest dwa razy lub więcej niż szerokość, nazywane są pałeczkami; jeśli różnica między długością a szerokością jest niewielka, nazywa się je bakteriami.
Bakterie o kręconych kształtach różnią się nie tylko długością i grubością, ale także liczbą i charakterem loków. Lekko zakrzywione bakterie (zakręcenie nie przekracza 1/4 obrotu spirali) nazywane są vibrios, bakterie, które mają jeden lub więcej dużych regularnych loków, nazywane są spirillum. Długie i cienkie bakterie o zawiłym kształcie z licznymi małymi lokami, czasami z dużymi krzywiznami całego włókna, nazywane są krętkami.
Struktura bakterii
Dzięki prostocie swojej organizacji i niewielkim rozmiarom bakterie należą do najbardziej elementarnych istot i stoją na najniższych szczeblach życia. Pomimo ogromnych postępów w nauce i technologii, nie wszystkie kwestie dotyczące budowy bakterii zostały jeszcze rozwiązane.
Ciało bakterii składa się ze skorupy i protoplazmy o etycznej zawartości, impregnowanych sokiem komórkowym. Powłoka bakterii jest cienka, bezbarwna, jej struktura nie jest widoczna pod mikroskopem. Aby to zobaczyć, uciekają się do sztucznych metod przetwarzania. Powłoka leży u podstaw zewnętrznego kształtu komórki i wydaje się być dobrze znaną obroną przed niekorzystnymi warunkami. Swobodnie otulając zawartość komórki, dzięki swojej elastyczności umożliwia swobodny ruch bakterii, któremu często towarzyszą energiczne ruchy całego ciała.
Zewnętrzne warstwy muszli, chłonące wodę, często pęcznieją i tworzą galaretowatą, lepką masę, osiągając zauważalne rozmiary. Gdy zewnętrzne warstwy stają się śluzowate, powłoka jest stale odnawiana kosztem protoplazmy. Schłodzona powłoka samoprzylepna nazywana jest kapsułką. Intensywność tworzenia śluzu i kapsułek zależy od właściwości żywienia i czasami może być bardzo znacząca. Kapsułka śluzowa powstaje nie tylko w pobliżu każdej komórki z osobna, ale także w wielu komórkach połączonych w jedną kolonię i zamkniętych niejako we wspólnej kapsułce. Te oślizgłe kolonie bakterii nazywają się zoogle. Tworzenie kapsułek nie jest charakterystyczne dla wszystkich rodzajów bakterii.
ruch bakterii
Zdolność do spontanicznego ruchu jest nieodłączna tylko w niektórych grupach bakterii. Bakterie poruszają się za pomocą wici lub rzęsek. Wici wyglądają jak mniej lub bardziej długie włókna. Są bardzo delikatne, cienkie, łatwo się łamią i nie są widoczne pod mikroskopem bez specjalnego zabarwienia. Ich średnica nie przekracza 1/20 średnicy ciała bakterii.
Mobilne formy bakterii dzielą się na następujące grupy:
- monotrichous - jest tylko jedna wici polarnej,
- lofotryczny - na jednym końcu komórki znajduje się wiązka wici,
- wici okostne znajdują się na całej powierzchni ciała.
Charakter umiejscowienia wici na ciele bakterii determinuje charakter jej ruchu - prostoliniowy lub losowy. Mobilność bakterii zależy od wielu warunków: temperatury, składu pożywki, produktów ich aktywności życiowej itp. Ruchliwe formy bakterii nie są wyposażone w wici na wszystkich etapach ich rozwoju i nie we wszystkich warunkach wzrostu.
zarodnikowanie
W ciele wielu bakterii, w pewnych okresach ich rozwoju, pojawiają się okrągłe lub elipsoidalne formacje - podpory. Zwykle kończą cykl rozwojowy bakterii. Rozmiar zarodników w porównaniu z rozmiarem komórek, które je wytworzyły, może się znacznie różnić.
Podpory nie tworzą się u wszystkich rodzajów bakterii. Otacza je dobrze odizolowana muszla, prawie nieprzepuszczalna dla wody i są najbardziej stabilnymi formacjami w całym świecie żywych. Dlatego często wytrzymują gotowanie przez kilka godzin i przedłużoną ekspozycję na suchą parę w temperaturach od 120 do 140 °. Zarodniki wielu pałeczek zachowują swoją żywotność po długim przebywaniu w temperaturze -190 °, a nawet w temperaturze ciekłego wodoru (-253 °). Są również odporne na działanie chemikaliów – trucizn. Wszystko to sprawia, że walka z chorobotwórczymi gatunkami zarodników bakterii jest niezwykle trudna.
Dojrzały zarodnik może zachować żywotność przez dziesięciolecia. Zwykle kiełkowanie zarodników następuje po pewnym okresie uśpienia i wiąże się z wpływem warunków zewnętrznych. Cały proces sporulacji następuje w ciągu jednego dnia lub mniej. Gdy zarodnik dojrzeje, komórka, która go wyprodukowała, stopniowo obumiera i pojawia się dojrzały zarodnik. Podczas kiełkowania pęcznieje, staje się bogatszy w wodę, z której wychodzi sadzonka ubrana w cienką skorupkę.
Reprodukcja bakterii
Po osiągnięciu stanu dojrzałości i limitu wzrostu bakterie zaczynają się rozmnażać przez prosty podział. Podczas podziału w środkowej części ciała bakterii pojawia się przegroda, która następnie dzieli i oddziela dwie nowe komórki. Kolejne rozmieszczenie przegród podczas podziału jest różne dla różnych bakterii. W formach pręcikowych znajduje się prostopadle do osi długiej, w formach kulistych przegrody mogą znajdować się w jednej, dwóch lub trzech wzajemnie prostopadłych płaszczyznach, co jest przyczyną powstawania takich form jak paciorkowce, tetrakoki i sarcyny .
Szybkość reprodukcji bakterii zależy od wielu warunków i może być bardzo różna. Im korzystniejsze są warunki zewnętrzne dla istnienia bakterii, tym szybciej następuje ich podział. W normalnych warunkach liczba bakterii podwaja się co około pół godziny. Gdyby zawsze działo się to bez przeszkód, to liczba bakterii z jednej komórki osiągnęłaby kolosalne proporcje. Według mikrobiologa Kohna potomstwo jednego Bacillus w ciągu pięciu dni może wypełnić wszystkie morza i oceany. Jednak to się nigdy nie wydarzyło i nigdy nie nastąpi. Cykl życiowy bakterii jest ograniczony pewnymi warunkami zewnętrznymi, po przekroczeniu których rozmnażanie ulega spowolnieniu lub całkowicie ustaje. Brak żywienia, szkodliwe produkty przemiany materii, konkurencja różnych gatunków itp. mają szkodliwy wpływ na bakterie. W niesprzyjających warunkach giną masowo.
Klasyfikacja bakterii
Pozycja bakterii w systemie istot żywych nie jest jeszcze dobrze określona. Powszechnie przyjmuje się, że bakterie są częścią świata roślin, a grzyby i glony są z nimi najbliżej spokrewnionymi organizmami. Cechy morfologiczne bakterii w większości przypadków ograniczają się do kilku form: kulistej, pręcikowej, spiralnej. Niezwykła prostota i elementarny charakter ich zewnętrznej organizacji utrudnia ich klasyfikację. Określenie gatunku bakterii na podstawie samych cech morfologicznych jest niemożliwe. Systematyka naukowa opiera się na morfologii i historii rozwoju, jednak do klasyfikacji bakterii niezbędna jest nie tylko znajomość morfologii, ale także ich właściwości fizjologicznych i biochemicznych. W związku z tym ustala się: stosunek bakterii do tlenu, warunki temperaturowe, tworzenie pigmentu, upłynnianie żelatyny, tworzenie kwasów i gazu na cukrach, zmianę mleka z rozwojem bakterii, powstawanie indolu, siarkowodoru, amoniaku, redukcja azotanów do azotynów lub do wolnego azotu. Nie zawsze to jednak wystarcza do określenia rodzaju bakterii.
Istnieją różne systemy klasyfikacji bakterii, ale wszystkie są warunkowe i znacznie mniej lub bardziej odbiegają od klasyfikacji naturalnej. Uwzględnienie tych systemów lub przynajmniej jednego z nich w tym przypadku nie jest konieczne nawet w przypadku zastosowania wobec bakterii fitopatogennych. Należy tylko powiedzieć, że obecnie prawie wszystkie fitopatogenne bakterie są zjednoczone w rodzajach Pseudomonas, Xanthomonas, Bacterium i Erwinia.
Ostatnio M. V. Gorlenko (1961) zaproponował następujący system klasyfikacji fitopatogennych bakterii z klasy Eubacteriales:
I. Rodzina Mycobacteriaceae(Chester, 1901) - bakterie nieruchome (bez wici):
- 1. rodzaj - Gorynebacterium (Leman i Neumann, 1896) - (bakterie gram-dodatnie;
- 2. rodzaj Aplanobacterium (Smith, 1905, Geshich, 1956) - bakterie Gram-ujemne.
II. Rodzina Pseudomonadaceae(Wilson i wsp., 1917) - bakterie z wiciami (polarne):
- 1. rodzaj - Pseudomonas (Migula, 1900) - bakterie niebarwione i fluorescencyjne;
- 2. rodzaj - Xanthomonas (Dawson, 1839) - bakterie z kolorowymi koloniami.
III. Rodzina Bacteriaceae(Kon, 1872) - ruchliwe bakterie z wiciami okostnymi, które nie tworzą podpór:
- 1. rodzaj - Bacterium (Ehrenberger, 1828) - formy niezabarwione, które nie tworzą pektynaz i protopektynaz;
- 2. rodzaj - Pectobacterium (Waldy, 1945) - niewybarwione formy tworzące pektynazę i protopektynazę;
- 3. rodzaj - Chromobacterium (Bergonzini, 1881) - formy kolorowe.
IV. Rodzina Bacillaceae(Fischer, 1895) - ruchliwe bakterie, pałeczki tworzące zarodniki:
- 1. rodzaj - Bacillus (Kon, 1832) - komórki nie pęcznieją ani nie pęcznieją nieznacznie podczas tworzenia zarodników;
- Drugi rodzaj - Clostridium (Praznovsky, 1880) - komórki pęcznieją podczas tworzenia zarodników.
W powyższym układzie pominięto dotychczas ogólnie przyjęty rodzaj Erwinia. Wyizolowano z niego specjalny rodzaj Pectobacterium, który obejmuje bakterie o działaniu pektolitycznym i okostnej wici. Te z bakterii fitopatogennych, które nie posiadają tej zdolności, przypisuje się do rodzaju Bacterium. System ten, sam w sobie racjonalny, jest nowy i nie wszedł jeszcze w życie codzienne, dlatego w tej pracy trzymamy się klasyfikacji, w której miejsce zajmuje rodzaj Erwinia. Ta rodzajowa nazwa bakterii fitopatogennych jest szeroko stosowana w literaturze specjalistycznej zarówno w naszym kraju, jak i za granicą.
Określenie rodzaju bakterii bez użycia sztucznych pożywek jest niemożliwe. W związku z tym można zauważyć, że podczas hodowli bakterii tworzą bardzo charakterystyczne kolonie. W takim przypadku do oceny gatunku bakterii można wykorzystać jeden wygląd.
Jeśli znajdziesz błąd, zaznacz fragment tekstu i kliknij Ctrl+Enter.
1.5. Współczesna klasyfikacja bakterii We współczesnej taksonomii bakterii rozwinęła się sytuacja charakterystyczna również dla klasyfikacji innych organizmów: poczyniono postępy w tworzeniu systemu klasyfikacji filogenetycznej, który odzwierciedla główne kierunki rozwoju ewolucyjnego i relacje przedstawicieli niektóre taksony, ale zjawiska sztuczne zachowują swoje znaczenie - typowe klasyfikacje, wygodniejsze do identyfikacji drobnoustrojów. Obecnie nie ma żadnego szczegółowego systemu ewolucyjnego prokariotów i najprawdopodobniej rozwiązanie tego problemu nie jest kwestią najbliższej przyszłości. Cechy prokariontów z zakresu organizacji morfologicznej, fizjologicznej, biochemicznej i genetycznej wskazują na nieprzydatność do nich dobrze rozwiniętych zasad stosowanych w budowie systemu organizmów wyższych. Nie zagłębiając się w historyczne aspekty problemu taksonomii bakterii, należy zauważyć, że najbardziej akceptowalnym systemem klasyfikacji filogenetycznej dla prokariotów jest system oparty na dopasowaniu sekwencji nukleotydów w 16S rRNA. System ten stał się podstawą drugiej edycji wielotomowej encyklopedii prokariontów - Bergey's Manual of Systematic Bacteriology (Bergey's Manual of Systematic Bacteriology), której pierwszy tom został opublikowany w 2001 roku. W tej pracy wszystkie prokarionty zostały podzielone na 26 filogenetycznych „gałęzi” (grup) opartych na strukturze ich 16S-rRNA; 23 „gałęzie” są reprezentowane przez eubakterie, a trzy przez archebakterie. Należy podkreślić, że duża liczba tych grup filogenetycznych zawiera gatunki prokariotyczne, które nie zostały wyizolowane jako czyste kultury i dlatego nie zostały jeszcze szczegółowo zbadane. W przypadku przedstawicieli tych gatunków znane są obecnie tylko sekwencje nukleotydowe 16S rRNA. Spośród 23 grup eubakterii dwie grupy filogenetyczne są reprezentowane przez bakterie Gram-dodatnie, pozostałe grupy są Gram-ujemne. Bakterie Gram-ujemne składają się z dużej grupy Proteobacteria (Proteobacteria) i 20 grup innych bakterii z tego typu ścianą komórkową. Krótki opis Proteobacteria, do których mitochondria i chloroplasty większości eukariontów są najbliższe w składzie 16S-rRNA, przedstawiono w Tabeli 1. 2. Proteobacteria to grupa bakterii Gram-ujemnych, która jest bardzo niejednorodna pod względem morfologicznym, fizjologicznym i biochemicznym. Przedstawiciele tej grupy charakteryzują się wszystkimi typami metabolizmu energetycznego i żywienia. Komórki większości gatunków Proteobacteria mają kształt pręcika, kuli lub wibrioidu, mnożą się głównie przez rozszczepienie binarne, ale niektóre gatunki charakteryzują się pączkowaniem i tworzeniem owocników w złożonym cyklu komórkowym. W tej grupie znajdują się zarówno bakterie ruchliwe z powodu wici, jak i nieruchome. W odniesieniu do tlenu cząsteczkowego Proteobacteria są klasyfikowane jako bezwzględne tlenowce, obligatoryjne i fakultatywne beztlenowce. Grupa Proteobacteria podzielona jest na pięć podgrup w oparciu o różnice w 16S rRNA: alfa, beta, gamma, delta i epsilon. Oprócz Proteobakterii, eubakterie Gram-ujemne obejmują następujące główne grupy eubakterii: termofile wodorowe, zielone bakterie nitkowate, zielone bakterie siarkowe, cyjanobakterie, krętki, cytofagi, bakteroidy, chlamydia, planctomycetes, deinococci, chloroflexus, fusobakterie, termobakterie, itp. Grupy filogenetyczne bakterii Gram-dodatnich - Actinobacteria i Firmicutes. Grupa Actinobacteria („gałąź promieniowców”) reprezentowana jest przez następujące rodzaje bakterii o wysokiej zawartości par GC w DNA: Geodermatophilus, Frankia, Streptomyces, Arthrobacter, Micrococcus, Actinomyces, Bifidobacterium, Propionibacterium, Actinoplanes, Nocardia, Rhodococcus, Corynebacterium , prątki. Grupa Firmicutes („gałąź Clostridium” - głównie bakterie Gram-dodatnie z niską zawartością par GC w DNA) obejmuje następujące rodzaje: Clostridium, Lactococcus, Pediococcus, Streptococcus, Enterococcus, Leuconostoc, Listeria, Caryophanon, Staphylococcus, Sarcina , Sporosarcina, Bacillus, Desulfotomaculum, Heliobacterium, Mycoplasma, Ureaplasma itp. Archaebakterie dzielą się na trzy grupy filogenetyczne: Crenarchaeota, Euryarchaeota i Korarchaeota. Grupa Crenarchaeota składa się z niezwykle ciepłolubnych bakterii, których większość przedstawicieli metabolizuje siarkę, niektórzy redukują jony żelaza i molibdenu. Grupa Euryarchaeota obejmuje obligatoryjne beztlenowe archebakterie metanogenne, a także ekstremalne termofile i halofile. Grupę Korarchaeota tworzą archebakterie żyjące w gorących źródłach siarkowych. Do tej pory żaden z przedstawicieli tej grupy (posiadający podobny 16S rRNA) nie został wyizolowany jako czysta kultura, dlatego ich cechy fenotypowe nie zostały wystarczająco zbadane. Kończąc rozważania na temat gałęzi filogenetycznych prokariotów, należy zauważyć, że proponowany system filogenetyczny, oparty na badaniu sekwencji nukleotydowych tylko jednego genu rybosomalnego RNA, jest niczym innym jak jednym z wygodnych technicznie i opracowanych systemów porządkowania licznych organizmów w celu ich identyfikacji, dlatego logiczne jest skonstruowanie prawidłowej taksonomii bakterii tylko przy uwzględnieniu tej cechy nie jest możliwe. Najbardziej rozpoznawalną i stosowaną klasyfikacją fenotypową bakterii jest ta przedstawiona w dziewiątym wydaniu Burgey's Guide to Bacteria. W tym wydaniu bakterie podzielono na cztery główne kategorie (podziały) w oparciu o strukturę warstwy granicznej komórki: 1) Gracilicutes (z łac. cuts - skóra, gracilis - cienka) - gram-ujemne eubakterie ze ścianami komórkowymi; 2) Firmicutes (od łac. firmus - silny) - gram-dodatnie eubakterie ze ścianami komórkowymi; 3) Tenericutes (od łacińskiego tener - miękki, delikatny) - eubakterie, pozbawione ścian komórkowych; 4) Mendosicutes (z łac. mendosus - błędne) - archebakterie, których ściany komórkowe różnią się od podobnych struktur innych prokariotów. Oddział Gracilicutes obejmuje bakterie o różnej morfologii ze ścianą komórkową Gram-ujemną. Rozmnażanie następuje głównie przez rozszczepienie binarne, niektóre bakterie rozmnażają się przez pączkowanie. Endospore nie tworzy się. Większość jest ruchliwa: istnieją wszystkie rodzaje ruchu bakterii - za pomocą wici, przesuwanie się, zginanie. Podział obejmuje bakterie tlenowe, beztlenowe i fakultatywne beztlenowe; bakterie fototroficzne i chemotroficzne. Oddział podzielony jest na trzy klasy: Scotobacteria, Oxyphotobacteria, Anoxyphotobacteria. Klasa Scotobacteria obejmuje bakterie Gram-ujemne, które nie wykorzystują energii świetlnej do celów metabolicznych, ale otrzymują ją tylko w wyniku reakcji redoks. Nazwa klasy pochodzi z greki scotos - ciemność. To największa klasa bakterii. Klasa Anoxyphotobacteria obejmuje bakterie purpurowe, bakterie zielone i heliobakterie, które przeprowadzają fotosyntezę anoksygeniczną (bez uwalniania tlenu cząsteczkowego). Klasa Oxyphotobacteria jest reprezentowana przez cyjanobakterie i prochlorofity, które przeprowadzają fotosyntezę tlenową (z uwalnianiem tlenu cząsteczkowego). Ten rodzaj fotosyntezy jest podobny do fotosyntezy zachodzącej w roślinach. Dział Firmicutes obejmuje bakterie ze ścianą komórkową Gram-dodatnią. Komórki mogą mieć różne kształty: pręciki, kokcy, nitkowate, rozgałęzione. Niektórzy przedstawiciele tworzą endospory. Większość z nich jest nieruchoma; formy ruchliwe mają wici okostne. Oddział obejmuje bakterie tlenowe, beztlenowe i fakultatywne beztlenowe. Oddział składa się z dwóch klas: Firmibacteria, Thallobakterie. Klasa Firmibacteria obejmuje dużą liczbę „nierozgałęzionych” bakterii Gram-dodatnich. Klasa Thallobacteria obejmuje bakterie, których komórki są zdolne do „rozgałęzienia”. Dział Tenericutes jest reprezentowany przez bakterie, które nie mają ściany komórkowej. Ze względu na brak ściany komórkowej kształt komórek nie jest stały: w czystej kulturze jednego gatunku jednocześnie obecne są komórki kokoidalne, pręcikowe, nitkowate, gruszkowate, w kształcie dysku i inne. Rozmnażanie bakterii zawartych w tej sekcji następuje przez rozszczepienie binarne, pączkowanie. Barwienie Grama jest ujemne. Charakterystyczne jest tworzenie się małych kolonii wyrastających w agar. Mogą być saprofityczne, pasożytnicze lub chorobotwórcze. Oddział składa się z jednej klasy Mollicutes (mykoplazm). Podział Mendosicutes tworzą bakterie o sztywnej ścianie komórkowej, ale pozbawione peptydoglikanu mureinowego. Większość przedstawicieli to ścisłe beztlenowce, z których wiele ma wici. Gatunki charakteryzują się różnorodnością ekologiczną i metaboliczną, zdolnością do życia w ekstremalnych warunkach. Oddział składa się z jednej klasy - Archaebacteria. W ramach czterech działów (kategorii głównych) zidentyfikowano 35 grup (lub sekcji) bakterii, które w większym lub mniejszym stopniu zostaną scharakteryzowane w kolejnych rozdziałach. Następujące grupy należą do działu Gracilicutes. Grupa 1. Krętki. Grupa 2. Tlenowe (lub mikroaerofilne), ruchliwe, spiralne (lub wibrioidalne) bakterie Gram-ujemne. Grupa 3. Nieruchliwe lub rzadko ruchliwe zakrzywione bakterie Gram-ujemne. Grupa 4. Gram-ujemne pałeczki i ziarniaki tlenowe (lub mikroaerofilne). Grupa 5. Opcjonalne aerobowe pałeczki Gram-ujemne. Grupa 6. Gram-ujemne beztlenowe pręciki proste, zakrzywione lub spiralne. Grupa 7. Bakterie przeprowadzające dysymilacyjną redukcję siarki lub siarczanu. Grupa 8. Beztlenowe ziarniaki Gram-ujemne. Grupa 9. Rickettsia i chlamydia. Grupa 10. Anoksygenne bakterie fototroficzne. Grupa 11. Tlenowe bakterie fototroficzne. Grupa 12. Tlenowe bakterie chemolitotroficzne i organizmy pokrewne. Grupa 13. Pączkujące i (lub) rozwijające się bakterie. Grupa 14. Bakterie z osłonkami. Grupa 15. Niefotosyntetyczne bakterie poślizgowe, które nie tworzą owocników. Grupa 16. Bakterie poślizgowe tworzące owocniki. Dział Firmicutes obejmuje: Grupa 17. Gram-dodatnie ziarniaki. Grupa 18. Gram-dodatnie pałeczki i ziarniaki tworzące endospory. Grupa 19. Gram-dodatnie pałeczki o prawidłowej formie, które nie tworzą zarodników. Grupa 20. Gram-dodatnie pałeczki o nieregularnym kształcie, które nie tworzą zarodników. Grupa 21. Mykobakterie. Grupy 22–29. Promieniowce. Do działu Tenericutes należą: Grupa 30. Mykoplazmy. Dział Mendosicutes obejmuje: Grupa 31. Metanogeny. Grupa 32. Archebakterie redukujące siarczany. Grupa 33. Skrajnie halofilne archebakterie (halobakterie). Grupa 34. Archebakterie bez ściany komórkowej. Grupa 35 Wyjątkowo termofilne i hipertermofilne archebakterie metabolizujące siarkę. Podsumowując należy podkreślić, że większość drobnoustrojów występujących w naturalnych zbiorowiskach nadal wymaga izolacji do czystych kultur. Uważa się, że obecnie można hodować tylko 0,1% całkowitej różnorodności drobnoustrojów, a nie ma możliwości wyhodowania i identyfikacji pozostałych przedstawicieli bakterii, chociaż około 5 tysięcy gatunków prokariotów zostało już wyizolowanych i opisanych w czystej kulturze.